
Что такое генетический код и какие его свойства

Хотя генетика — невероятно сложная наука, генетический код как раз довольно прост для понимания, его изучают в девятом классе на уроках биологии и потом благополучно забывают.
Что такое генетический код?
Генетический код — это правила, по которым последовательность аминокислот в белке кодируется молекулой матричной РНК.
Белок — это молекула, состоящая из цепочки аминокислот, соединенных пептидной связью. Всего известно около 60 аминокислот, но в большинстве белков используются 20 так называемых стандартных аминокислот.
Свойства белков зависят от того, какие аминокислоты, в каком количестве и в какой последовательности входят в их состав. Скажем, антитела к различным вирусам, инсулин, альбумин (яичный белок), гемоглобин, многие ферменты, основа клеточной мембраны — все это белки, но очень разные.
ДНК и РНК — молекулы, состоящие из цепочки нуклеотидов (нуклеиновых кислот). Всего существует пять нуклеотидов: аденин, гуанин, цитозин,тимин и урацил. Первые три присутствуют и в ДНК и в РНК. Тимин и урацил — это похожие по составу нуклеотиды, тимин присутствует только в ДНК, урацил — только в РНК.
В ДНК последовательностью нуклеотидов зашифрована генетическая информация о составе всех белков, необходимых конкретному организму. РНК эту информацию считывает и передает “инструкцию” непосредственно на место синтеза белка, в рибосому. В каждой из примерно 10 млрд человеческих клеток постоянно работает заводик по производству белков — без перерыва и отпуска.
Схема в общих чертах выглядит так: в ядре клетки постоянно формируется слепок ДНК — молекула матричной РНК. Каждому из четырех нуклеотидов-”кирпичиков” ДНК соответствует определенный нуклеотид мРНК — они комплементарны, то есть дополняют друг друга, как ключ и замок. Аденин соединяется с урациломтимином, цитозин с гуанином. Нуклеотиды обозначаются первыми буквами названий, чаще латинскими. Процесс создания “слепка” называется транскрипцией. Он происходит непрерывно.
Готовая молекула матричной РНК отделяется от ДНК и попадает из ядра в рибосому. Молекулы транспортной РНК тащат к ней, как к конвейеру, необходимые аминокислоты. Эти аминокислоты в рибосоме соединяются в цепочку по инструкции, считанной с матричной РНК: каждую аминокислоту кодирует тройка (триплет) нуклеотидов мРНК, стоящих в определенном порядке (скажем, GGU, AGU, CUA и т.д). Процесс сборки белка называется трансляцией.
Соответствие между триплетами нуклеотидов и конкретными аминокислотами — это генетический код.
Свойства генетического кода
Практически всегда и у всех организмов на Земле соответствие между триплетами нуклеотидов и конкретными аминокислотами одинаковое — это свойство называют универсальностью генетического кода. Генетический код универсален для всех, от цианобактерий до человека. Это лишний раз напоминает о том, что мы с цианобактериями хоть и далекие, но родственники.
Однако в некоторых случаях генетический код все-таки немного видоизменяется, в частности, при синтезе белков в митохондриях.
Специфичность генетического кода состоит в том, что каждому триплету-кодону соответствует только одна аминокислота. Например, триплет AUG кодирует только аминокислоту метионин; триплет CAC — только гистидин; UAU — только тирозин.
Однако вариантов тройных комбинаций из четырех нуклеотидов существует 43 = 64. А аминокислот в белках — всего 20. Поэтому природа устроила так, что каждую аминокислоту кодируют несколько триплетов. Скажем, аминокислоту аланин можно закодировать четырьмя кодонами: GCU, GCA, GCC, GCG (все начинаются с GC). А лейцин — аж семью кодонами, из них четыре начинаются с CU. Это свойство называется избыточностью генетического кода. Оно выполняет важную роль: предупреждает ошибки и поломки в системе. Когда в ДНК есть мутация, матричная РНК тоже копирует эту ошибку. Но если мутация точечная, то есть лишь один нуклеотид в триплете неправильный, то благодаря избыточности генетического кода есть шанс, что триплет все равно закодирует нужную аминокислоту.
Сейчас установлены многие мутации, приводящие к наследственным заболеваниям. Например, из-за перестройки в Х-хромосоме в организме перестает вырабатываться белок под названием «фактор VIII». Этот белок необходим для сворачивания крови и формирования сгустка на ране. Болезнь известна как гемофилия. Более мелкие, точечные мутации приводят к талассемии, серповидной анемии, муковисцидозу и другим наследственным заболеваниям.
Есть и другие болезни, о наследственном происхождении которых генетики пока только строят предположения, — например, синдром Туретта. Возможно, в будущем удастся найти мутацию или, скорее, набор мутаций, ответственных за его возникновение.
Непрерывность генетического кода
Между отдельными триплетами матричной РНК никакого разграничения нет. Поэтому в синтезе белка окончание одного триплета автоматически означает начало следующего. Отсюда еще два свойства генетического кода: непрерывность и неперекрываемость. Непрерывность генетического кода — это считывание кодонов без промежутков, один за другим. Неперекрываемость генетического кода означает, что каждый нуклеотид может входить в состав только одного триплета. Если триплеты GUA и CCG выстроены в непрерывную цепочку, то A не может быть частью триплета ACC — поскольку триплеты не перекрываются.
Помимо смысловых кодонов, существуют кодоны, которые означают начало и конец белковой молекулы: UAA, UAG, UGA. В огромном большинстве случаев эти кодоны служат только сигнальными флажками, знаками препинания в закодированной фразе. Но есть исключения: в митохондриях, а также у инфузорий и зеленых водорослей триплеты UAA, UAG, UGA могут служить еще и смысловыми кодонами. Как рибосома “понимает” разницу? Этот вопрос не до конца решен, но считается, что значение стоп-кодонов зависит от положения в молекуле мРНК: если они попадаются в середине, значит, смысловые; если близко к концу молекулы РНК — рибосома воспринимает их как стоп-кодоны.
Таблица генетического кода
Благодаря таким жестким закономерностям генетический код легко складывается в таблицу.
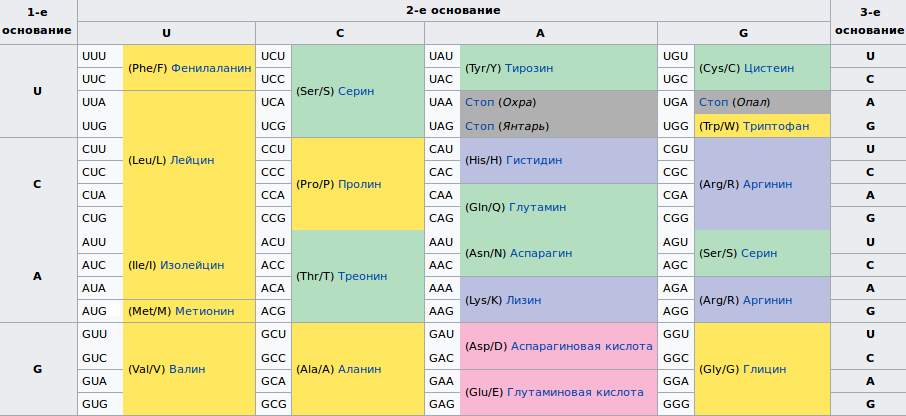
Расшифровка генетического кода
Как только в 1953 году Крик и Уотсон на основании данных Франклин и Уилкинса открыли структуру двойной спирали ДНК, ученые задались вопросом, как именно ДНК кодирует последовательность аминокислотніх остатков в белке. Первым предложил идею триплетов Георгий Гамов, американский физик русского происхождения. Гамов не занимался биологией — он просто заметил, что для кодирования 20 аминокислот достаточно 64 комбинаций, возникающих при сочетании четырех нуклеотидов тройками.
В 1961 году эту гениальную в своей элегантной простоте идею подтвердили экспериментально. Американский биохимик Маршалл Ниренберг с коллегами провел эксперимент по синтезу белка (точнее, пептида) в пробирке, in vitro. В качестве матрицы взяли синтетический олигонуклеотид — нуклеиновую кислоту, состоящую только из урацила, и поместили ее в “подопытного кролика” — кишечную палочку E. Coli. Бактерия начала прилежно синтезировать белок на искусственной матрице. В результате получился полипептид, содержавший только аминокислоту фенилаланин. Так впервые было установлено значение триплета-кодона: кодон UUU кодирует фенилаланин.
Дальнейшие эксперименты проводились с синтетической РНК, содержащей только аденин, только цитозин, а затем — разные комбинации нклеотидов. Большинство кодонов расшифровал индийский биохимик Хар Гобинд Корана. А американский биохимик Роберт Холли описал структуру матричной РНК. В 1968 году эти трое ученых — Ниренберг, Корана и Холли — получили Нобелевскую премию по физиологии и медицине за вклад в расшифровку генетического кода.
Читайте также:
Генетики расшифровали геном саблезубой кошки
Анализ ДНК показал, что коренных жителей Карибского региона уничтожили еще до Колумба

Источник
ГЕНЕТИЧЕСКИЙ КОД (греч, genetikos относящийся к происхождению; син.: код, биологический код, аминокислотный код, белковый код, код нуклеиновых к-т) — система записи наследственной информации в молекулах нуклеиновых кислот животных, растений, бактерий и вирусов чередованием последовательности нуклеотидов.
Схема передачи наследственной информации (сплошными стрелками обозначены пути передачи информации): на молекуле ДНК осуществляется построение трех типов РНК: рРНК рибосомной (растворимой), иРНК — информационной, тРНК— транспортной, участвующих в синтезе белка и белковых компонентов прочих веществ клетки (пунктирными стрелками показано участие отдельных компонентов клетки в регуляции передачи информации); в процессе размножения образуется копия ДНК (ДНК1), для которой передача наследственной информации и ее регуляция идентичны данной схеме.
Генетическая информация (рис.) из клетки в клетку, из поколения в поколение, за исключением РНК-содержащих вирусов, передается путем редупликации молекул ДНК (см. Репликация). Реализация наследственной информации ДНК в процессе жизнедеятельности клетки осуществляется через 3 типа РНК: информационную (иРНК или мРНК), рибосомную (рРНК) и транспортную (тРНК), которые с помощью фермента РНК-полимеразы синтезируются на ДНК как на матрице. При этом последовательность нуклеотидов в молекуле ДНК однозначно определяет последовательность нуклеотидов во всех трех типах РНК (см. Транскрипция). Информацию гена (см.), кодирующего белковую молекулу, несет только иРНК. Конечным продуктом реализации наследственной информации является синтез белковых молекул, специфичность которых определяется последовательностью входящих в них аминокислот (см. Трансляция).
Поскольку в составе ДНК или РНК представлено только по 4 разных азотистых основания [в ДНК — аденин (А), тимин (Т), гуанин (Г), цитозин (Ц); в РНК — аденин (А), урацил (У), цитозин (Ц), гуанин (Г)], последовательность которых определяет последовательность 20 аминокислот в составе белка, возникает проблема Г. к., т. е. проблема перевода 4-буквенного алфавита нуклеиновых к-т в 20-буквенный алфавит полипептидов.
Впервые идея матричного синтеза белковых молекул с правильным предсказанием свойств гипотетической матрицы была сформулирована Н. К. Кольцовым в 1928 г. В 1944 г. Эйвери (О. Avery) с соавт, установил, что за передачу наследственных признаков при трансформации у пневмококков ответственны молекулы ДНК. В 1948 г. Чаргафф (E. Chargaff) показал, что во всех молекулах ДНК имеет место количественное равенство соответствующих нуклеотидов (А-T, Г-Ц). В 1953 г. Ф. Крик, Дж. Уотсон и Уилкинс (М. H. F. Wilkins), исходя из этого правила и данных рентгеноструктурного анализа (см.), пришли к выводу, что молекул а ДНК представляет собой двойную спираль, состоящую из двух полинуклеотидных нитей, соединенных между собой водородными связями. Причем против А одной цепи во второй может находиться только Т, против Г — только Ц. Эта комплементарность приводит к тому, что последовательность нуклеотидов одной цепи однозначно определяет последовательность другой. Второй существенный вывод, вытекающий из этой модели,— молекула ДНК способна к самовоспроизведению.
В 1954 г. Гамов (G. Gamow) сформулировал проблему Г. к. в ее современном виде. В 1957 г. Ф. Крик высказал Гипотезу адаптера, предположив, что аминокислоты взаимодействуют с нуклеиновой к-той не непосредственно, а через посредников (теперь известных под названием тРНК). В ближайшие после этого годы все принципиальные звенья общей схемы передачи генетической информации, вначале гипотетичные, были подтверждены экспериментально. В 1957 г. были открыты иРНК [А. С. Спирин, А. Н. Белозерский с соавт.; Фолькин и Астрахан (E. Volkin, L. Astrachan)] и тРНК [Хоугленд (М. В. Hoagland)]; в 1960 г. синтезирована ДНК вне клетки с использованием в качестве матрицы существующих макромолекул ДНК (А. Корнберг) и открыт ДНК-зависимый синтез РНК [Вейсс (S. В. Weiss) с соавт.]. В 1961 г. была создана бесклеточная система, в к-рой в присутствии естественной РНК или синтетических полирибонуклеотидов осуществлялся синтез белковоподобных веществ [М. Ниренберг и Маттеи (J. H. Matthaei)]. Проблема познания Г. к. состояла из исследования общих свойств кода и собственно его расшифровки, т. е. выяснения, какие комбинации нуклеотидов (кодоны) кодируют определенные аминокислоты.
Общие свойства кода были выяснены независимо от его расшифровки и в основном до нее путем анализа молекулярных закономерностей образования мутаций (Ф. Крик и соавт., 1961; Н. В. Лучник, 1963). Они сводятся к следующему:
1. Код универсален, т. е. идентичен, по крайней мере в основном, для всех живых существ.
2. Код триплетен, т. е. каждая аминокислота кодируется тройкой нуклеотидов.
3. Код неперекрывающийся, т. е. данный нуклеотид не может входить в состав более чем одного кодона.
4. Код вырожден, т. е. одна аминокислота может кодироваться несколькими триплетами.
5. Информация о первичной структуре белка считывается с иРНК последовательно, начиная с фиксированной точки.
6. Большинство возможных триплетов имеет «смысл», т. е. кодирует аминокислоты.
7. Из трех «букв» кодона преимущественное значение имеют лишь две (облигатные), третья же (факультативная) несет значительно меньшую информацию.
Прямая расшифровка кода состояла бы в сравнении последовательности нуклеотидов в структурном гене (или синтезированной на нем иРНК) с последовательностью аминокислот в соответствующем белке. Однако такой путь пока технически невозможен. Были применены два других пути: синтез белка в бесклеточной системе с использованием в качестве матрицы искусственных полирибонуклеотидов известного состава и анализ молекулярных закономерностей образования мутаций (см.). Первый принес положительные результаты раньше и исторически сыграл в расшифровке Г. к. большую роль.
В 1961 г. М. Ниренберг и Маттеи применили в качестве матрицы гомо-полимер — синтетическую полиуридиловую к-ту (т. е. искусственную РНК состава УУУУ…) и получили полифенилаланин. Из этого следовало, что кодон фенилаланина состоит из нескольких У, т. е. в случае триплетного кода расшифровывается как УУУ. Позже наряду с гомополимерами были использованы полирибонуклеотиды, состоявшие из разных нуклеотидов. При этом был известен только состав полимеров, расположение же нуклеотидов в них было статистическим, поэтому и анализ результатов был статистическим и давал косвенные выводы. Довольно быстро удалось найти хотя бы по одному триплету для всех 20 аминокислот. Выяснилось, что присутствие органических растворителей, изменение pH или температуры, некоторые катионы и особенно антибиотики делают код неоднозначным: те же кодоны начинают стимулировать включение других аминокислот, в некоторых случаях один кодон начинал кодировать до четырех разных аминокислот. Стрептомицин влиял на считывание информации как в бесклеточных системах, так и in vivo, причем был эффективен только на стрептомицинчувствительных штаммах бактерий. У стрептомицинзависимых штаммов он «исправлял» считывание с кодонов, изменившихся в результате мутации. Подобные результаты давали основание сомневаться в правильности расшифровки Г. к. с помощью бесклеточной системы; требовалось подтверждение, и в первую очередь данными in vivo.
Основные данные о Г. к. in vivo получены при анализе аминокислотного состава белков у организмов, обработанных мутагенами (см.) с известным механизмом действия, напр, азотистой к-той, к-рая вызывает в молекуле ДНК замену Ц на У и А на Г. Полезную информацию дают также анализ мутаций, вызванных неспецифическими мутагенами, сравнение различий в первичной структуре родственных белков у разных видов, корреляция между составом ДНК и белков и т. п.
Расшифровка Г. к. на основании данных in vivo и in vitro дала совпадающие результаты. Позже были разработаны три других метода расшифровки кода в бесклеточных системах: связывание аминоацил-тРНК (т. е. тРНК с присоединенной активированной аминокислотой) тринуклеотидами известного состава (М. Ниренберг и соавт., 1965), связывание аминоацил-тРНК полинуклеотидами, начинающимися с определенного триплета (Маттеи с соавт., 1966), и использование в качестве иРНК полимеров, в которых известен не только состав, но и порядок нуклеотидов (X. Корана и соавт., 1965). Все три метода дополняют друг друга, а результаты находятся в соответствии с данными, полученными в опытах in vivo.
В 70-х гг. 20 в. появились методы особенно надежной проверки результатов расшифровки Г. к. Известно, что мутации, возникающие под действием профлавина, состоят в выпадении или вставке отдельных нуклеотидов, что приводит к сдвигу рамки считывания. У фага Т4 был вызван профлавином ряд мутаций, при которых изменился состав лизоцима. Этот состав был проанализирован и сопоставлен с теми кодонами, которые должны были получиться при сдвиге рамки считывания. Получилось полное соответствие. Дополнительно этот метод позволил установить, какие именно триплеты вырожденного кода кодируют каждую из аминокислот. В 1970 г. Адамсу (J. М. Adams) с сотрудниками удалось провести частичную расшифровку Г. к. прямым методом: у фага R17 определили последовательность оснований во фрагменте длиной в 57 нуклеотидов и сравнили с аминокислотной последовательностью белка его оболочки. Результаты полностью совпали с полученными менее прямыми методами. Т. о., код расшифрован полностью и верно.
Результаты расшифровки сведены в таблицу. В ней указан состав кодонов и РНК. Состав антикодонов тРНК комплементарен кодонам иРНК, т. е. вместо У в них находится А, вместо А — У, вместо Ц — Г и вместо Г — Ц, и соответствует кодонам структурного гена (той нити ДНК, с к-рой считывается информация) с той лишь разницей, что место тимина занимает урацил. Из 64 триплетов, которые могут быть образованы сочетанием 4 нуклеотидов, 61 имеет «смысл», т. е. кодирует аминокислоты, а 3 являются «нонсенсами» (лишенными смысла). Между составом триплетов и их смыслом имеется довольно четкая зависимость, к-рая была обнаружена еще при анализе общих свойств кода. В ряде случаев триплеты, кодирующие определенную аминокислоту (напр., пролин, аланин), характеризуются тем, что два первых нуклеотида (облигатные) у них одинаковы, а третий (факультативный) может быть любым. В других случаях (при кодировании, напр., аспарагина, глутамина) один и тот же смысл имеют два сходных триплета, у которых совпадают два первых нуклеотида, а на месте третьего стоит любой пурин или любой пиримидин.
Нонсенс-кодоны, 2 из которых имеют специальные названия, соответствующие обозначению фаговых мутантов (УАА-охра, УАГ-амбер, УГА-опал), хотя и не кодируют каких-либо аминокислот, но имеют большое значение при считывании информации, кодируя конец полипептидной цепи.
Считывание информации происходит в направлении от 51 -> 31 – к концу нуклеотидной цепи (см. Дезоксирибонуклеиновые кислоты). При этом синтез белка идет от аминокислоты со свободной аминогруппой к аминокислоте со свободной карбоксильной группой. Начало синтеза кодируется триплетами АУГ и ГУГ, которые в этом случае включают специфичную стартовую аминоацил-тРНК, а именно N-формилметио-нил-тРНК. Эти же триплеты при локализации внутри цепи кодируют соответственно метионин и валин. Неоднозначность снимается тем, что началу считывания предшествует нонсенс. Есть данные, говорящие в пользу того, что граница между участками иРНК, кодирующими разные белки, состоит более чем из двух триплетов и что в этих местах меняется вторичная структура РНК; этот вопрос находится в стадии исследования. Если нонсенс-кодон возникает внутри структурного гена, то соответствующий белок строится только до места расположения этого кодона.
Открытие и расшифровка генетического кода — выдающееся достижение молекулярной биологии — оказало влияние на все биол, науки, положив в ряде случаев начало развитию специальных крупных разделов (см. Молекулярная генетика). Эффект открытия Г. к. и связанных с ним исследований сравнивают с тем эффектом, который оказала на биол, науки теория Дарвина.
Универсальность Г. к. является прямым доказательством универсальности основных молекулярных механизмов жизни у всех представителей органического мира. Между тем большие различия в функциях генетического аппарата и его строении при переходе от прокариотов к эукариотам и от одноклеточных к многоклеточным, вероятно, связаны и с молекулярными различиями, исследование которых — одна из задач будущего. Поскольку исследования Г. к.— дело лишь последних лет, значение полученных результатов для практической медицины носит лишь Косвенный характер, позволяя пока понять природу заболеваний, механизм действия возбудителей болезней и лекарственных веществ. Однако открытие таких явлений, как трансформация (см.), трансдукция (см.), супрессия (см.), указывает на принципиальную возможность исправления патологически измененной наследственной информации или ее коррекции — так наз. генная инженерия (см.).
Таблица. ГЕНЕТИЧЕСКИЙ КОД
Первый нуклеотид кодона | Второй нуклеотид кодона | Третий , нуклеотид кодона | |||||||
У | Ц | А | Г | ||||||
У Урацил | УУУ | Фенилаланин | УЦУ | Серин | УАУ | Тирозин | УГУ | Цистеин | У |
УУЦ | УЦЦ | УАЦ | УГЦ | ц | |||||
УУА | Лейцин | УЦА | УАА* | J Нонсенс | УГА* | Нонсенс | А | ||
УУГ | УЦГ | УАГ* | УГГ | Триптофан | Г | ||||
Ц Цитозин | ЦУУ | Лейцин | ЦЦУ | Пролин | ЦАУ | Гистидин | ЦГУ | Аргинин | У |
ЦУЦ | ЦЦЦ | ЦАЦ J | ЦГЦ | ц | |||||
ЦУА | ЦЦА | ЦАА | Глутаминовая кислота | ЦГА | А | ||||
ЦУГ | ЦЦГ | ЦАГ | ЦГГ | Г | |||||
А Аденин | АУУ | Изолейцин | АЦУ | Треонин | ААУ | Аспарагиновая кислота | АГУ | Серин | У |
АУЦ | АЦЦ | ААЦ | АГЦ | Ц | |||||
АУА | АЦА | ААА | Лизин | АГА | Аргинин | А | |||
АУГ** | Метионин | АЦГ | ААГ | АГГ | Г | ||||
Г Гуанин | ГУУ | Валин | ГЦУ | Аланин | ГАУ | Аспарагин | ГГУ | Глицин | У |
ГУЦ | ГЦЦ | ГАЦ | ГГЦ | ц | |||||
ГУА | ГЦА | ГАА | Глутамин | ГГА | А | ||||
ГУГ** | Валин | ГЦГ | ГАГ | ГГГ | Г | ||||
* Кодирует конец цепи.
** Кодирует также начало цепи.
Библиография: Ичас М. Биологический код, пер. с англ., М., 1971; Лучник Н.Б. Биофизика цитогенетических поражений и генетический код, Л., 1968; Молекулярная генетика, пер. с англ., под ред. А. Н. Белозерского, ч. 1, М., 1964; Нуклеиновые кислоты, пер. с англ., под ред. А. Н. Белозерского, М., 1965; Уотсон Дж. Д. Молекулярная биология гена, пер. с англ., М., 1967; Физиологическая генетика, под ред. М. Е. Лобашева С. Г., Инге-Вечтомо-ва, Л., 1976, библиогр.; Desoxyribonuc-leins&ure, Schlttssel des Lebens, hrsg. v„ E. Geissler, B., 1972; The genetic code, Gold Spr. Harb. Symp. quant. Biol., v. 31, 1966; W o e s e C. R. The genetic code, N. Y. a. o., 1967.
H. B. Лучник.
Источник