
Какие продукты и сколько молекул атф получаются при гликолизе

Синтез АТФ при расщеплении глюкозы. Выделение энергии из гликогена
Мы можем определить общее количество молекул АТФ, которое образуется при расщеплении 1 молекулы глюкозы при оптимальных условиях.
1. Во время гликолиза образуются 4 молекулы АТФ: 2 молекулы АТФ расходуются на первом этапе фосфорилирования глюкозы, необходимого для хода процесса гликолиза, чистый выход АТФ при гликолизе равен 2 молекулам АТФ.
2. В итоге цикла лимонной кислоты образуется 1 молекула АТФ. Однако в связи с тем, что 1 молекула глюкозы расщепляется на 2 молекулы пировиноградной кислоты, каждая из которых проходит оборот в цикле Кребса, получается чистый выход АТФ на 1 молекулу глюкозы, равный 2 молекулам АТФ.
3. При полном окислении глюкозы суммарно образуются 24 атома водорода в связи с процессом гликолиза и циклом лимонной кислоты, 20 из них окисляются в соответствии с хемо-осмотическим механизмом с выделением 3 молекул АТФ на каждые 2 атома водорода. В итоге получается еще 30 молекул АТФ.
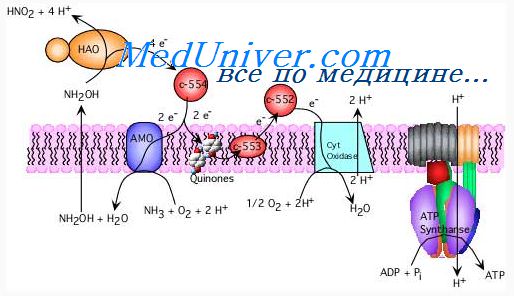
4. Четыре оставшихся атома водорода выделяются под влиянием дегидрогеназ и включаются в цикл хемоосмотического окисления в митохондриях помимо первой стадии. Окисление 2 атомов водорода сопровождается получением 2 молекул АТФ, в итоге получается еще 4 молекулы АТФ.
Сложив все полученные молекулы, получим 38 молекул АТФ как максимально возможное количество при окислении 1 молекулы глюкозы до углекислого газа и воды. Следовательно, 456000 калорий могут сохраняться в виде АТФ из 686000 калорий, получаемых при полном окислении 1 грамм-молекулы глюкозы. Эффективность преобразования энергии, обеспечиваемая этим механизмом, составляет около 66%. Остальные 34% энергии преобразуются в тепловую и не могут быть использованы клетками для выполнения специфических функций.
Выделение энергии из гликогена
Продолжительное высвобождение энергии из глюкозы, когда клетки не нуждаются в энергии, было бы слишком расточительным процессом. Гликолиз и последующее окисление атомов водорода постоянно контролируются в соответствии с потребностями клеток в АТФ. Этот контроль осуществляется многочисленными вариантами управляющих механизмов обратной связи в ходе химических реакций. К числу наиболее важных влияний такого рода можно отнести концентрацию АДФ и АТФ, контролирующую скорость химических реакций в ходе процессов обмена энергии.
Одним из важных путей, позволяющих АТФ управлять обменом энергии, является ингибирование фермента фосфофруктокиназы. Этот фермент обеспечивает образование фруктозо-1,6-дифосфата — одной из начальных стадий гликолиза, поэтому результирующим влиянием избытка АТФ в клетке будет торможение или даже остановка гликолиза, что, в свою очередь, приведет к торможению обмена углеводов. АДФ (равно как и АМФ) оказывает противоположное влияние на фосфофруктокиназу, существенно повышая ее активность. Когда АТФ используется тканями для энергообеспечения большинства химических реакций в клетках, это уменьшает ингибирование фермента фосфофруктокиназы, более того, его активность повышается параллельно увеличению концентрации АДФ. В результате запускаются процессы гликолиза, приводящие к восстановлению запасов АТФ в клетках.
Другой способ управления опосредован цитратами, образующимися в цикле лимонной кислоты. Избыток этих ионов существенно снижает активность фосфофруктокиназы, что не дает гликолизу опережать скорость использования пировиноградной кислоты, образующейся в результате гликолиза в цикле лимонной кислоты.
Третий способ, с помощью которого система АТФ-АДФ-АМФ может контролировать обмен углеводов и управлять выделением энергии из жиров и белков, заключается в следующем. Возвращаясь к различным химическим реакциям, служащим способом выделения энергии, мы можем заметить, что если весь имеющийся в наличии АМФ уже превращен в АТФ, дальнейшее образование АТФ становится невозможным. В результате прекращаются все процессы использования питательных веществ (глюкозы, белков и жиров) для получения энергии в виде АТФ. Лишь после использования образовавшегося АТФ в качестве источника энергии в клетках для обеспечения разнообразных физиологических функций вновь появляющиеся АДФ и АМФ запустят процессы получения энергии, в ходе которых АДФ и АМФ преобразуются в АТФ. Этот путь автоматически поддерживает сохранение определенных запасов АТФ, кроме случаев экстремальной активности клеток, например при тяжелых физических нагрузках.
– Также рекомендуем “Анаэробный гликолиз. Молочная и пировиноградная кислота”
Оглавление темы “Синтез АТФ. Обмен глюкозы и жиров”:
1. Образование АТФ через хемоосмотический механизм. Образование и синтез АТФ
2. Синтез АТФ при расщеплении глюкозы. Выделение энергии из гликогена
3. Анаэробный гликолиз. Молочная и пировиноградная кислота
4. Высвобождение энергии из глюкозы через пентозофосфатный цикл. Превращение глюкозы в жиры
5. Образование углеводов из белков и жиров. Регуляция глюконеогенеза
6. Метаболизм жиров в организме. Транспорт липидов
7. Свободные жирные кислоты. Транспорт свободных жирных кислот
8. Липопротеины. Виды и физиология липопротеинов
9. Обмен жиров и их депонирование. Жиры печени
10. Расщепление жирных кислот. Окисление жирных кислот
Источник
Гликолиз – процесс окисления глюкозы, при котором из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты, не является мембранозависимым процессом. Он происходит в цитоплазме. Однако ферменты гликолиза связаны со структурами цитоскелета. Суть гликолиза состоит в том, что молекула глюкозы (C6H12O6) без участия кислорода распадается на две молекулы пировиноградной кислоты (СН3СОСООН). При этом окисление идет за счет отщепления от молекулы глюкозы четырех атомов водорода, связывающихся со сложным органическим веществом НАД с получением двух молекул НАД•Н. Выделяющаяся при этом энергия запасается (40% от общего количества) в виде макроэргических связей двух молекул АТФ. 60% энергии выделяется в виде тепла. При последующем окислении НАД•Н получается еще 6 молекул АТФ. Таким образом, полный энергетический выход гликолиза в анаэробных условиях составляет 8 молекул АТФ.
На схеме в рамках обозначены исходные субстраты и конечные продукты гликолиза, цифрами в скобках – число молекул.
Для распада и частичного окисления молекулы глюкозы требуется протекание 11 сложных последовательных реакций.
Реакции гликолиза
Ход реакций
Ферменты, Активаторы, ингибиторы
Подготовительная стадия гликолиза
Стадия активации глюкозы проходит в 5 реакций, в ходе которых 1 молекула гексозы (глюкозы) расщепляется на 2 молекулы триоз-глицеральдегидфосфата
1. Необратимая реакция фосфорилирования глюкозы
Процесс гликолиза начинается с фосфорилирования глюкозы за счет АТФ – первая реакция. Это первая пусковая реакция гликолиза. Ее результатом является глюкозо-6-фосфат, имеющий отрицательный заряд. В гликолизе может участвовать не только глюкоза, но и другие гексозы (фруктоза), но в результате фосфорилирования и активации все равно образуется глюкозо-6-фосфат.

фермент: гексокиназа
Активаторы: АДФ, Н3РO4.
Ингибиторы: глюкозо-6-Ф, фосфоенолпируват.
2. Обратимая реакция изомеризации глюкозо-6-фосфата
Во второй реакции происходит изомеризация (внутримолекулярные перестройки) глюкозо-6-фосфата во фруктозо-6-фосфат.

фермент: глюкозо-6-фосфатизомераза
3. Необратимая реакция фосфорилирования фруктозо-6-фосфата (ключевая стадия гликолиза)
В третьей реакции происходит фосфорилирование (присоединение остатка ортофосфорной кислоты) фруктозо-6-фосфата с образованием фруктозо-1,6-дифосфата. При этом затрачивается еще одна молекула АТФ (уже вторая) – это вторая пусковая реакция гликолиза. Она идет в присутствии Mg2+ и является необратимой, так как сопровождается масштабным уменьшением свободной энергии.

фермент: фосфофруктокиназа
Активаторы: АДФ, АМФ, Н3РO4, К+.
Ингибиторы: АТФ, цитрат, НАДН.
4. Обратимая реакция дихотомического расщепления фруктозо-1,6-дифосфата
В четвертой реакции гликолиза происходит расщепление фруктозо-1,6-дифосфата на две молекулы глицеральдегид-3-фосфата.

фермент: алъдолаза
5. Обратимая реакция изомеризации дигидроксиацетона-3-фосфат в глицеральдегид-3-фосфат
В пятой реакции происходит изомеризация полученных триозофосфатов. На этом заканчивается первая стадия гликолиза.

фермент: триозофосфатизомераза
Стадия генерации АТФ
Проходит в 6 реакций (или 5), в ходе которых энергия окислительных реакций трансформируется в химическую энергию АТФ (субстратное фосфорилирование).
6. Окисление глицеральдегид-3-фосфата до 1,3-дифосфоглицерата (реакция гликолитической оксиредукции)
В шестой реакции происходит окисление альдегидной группы до карбоксильной. Выделившийся Н+ акцептируется NAD, который восстанавливается до NADH. Освобождающаяся энергия затрачивается для образования высокоэнергетической связи 1,3-бифосфоглицерата (1,3-бифосфоглицериновая кислота).

фермент: глицералъдегид-3-фосфат-дегидрогеназа
7. Субстратное фосфорилирование АДФ (7)
В седьмой реакции фосфорильная группа 1,3-бифосфоглицерата переносится на ADP, в результате чего образуется АТР (напоминаем, что следует иметь в виду две параллельные цепи реакций, с участием двух молекул триоз, образовавшихся из одной молекулы гексозы, следовательно, синтезируется не одна, а две молекулы АТР).

фермент: фосфоглицераткиназа
8. Реакция изомеризации 3-фосфоглицерата в 2-фосфоглицерат
В восьмой реакции гликолиза происходит перенос фосфатной группы с третьего атома углерода на второй. В результате образуется 2-фосфоглицерат (2-фосфоглицериновая кислота).

9. Реакция енолизации
Девятая реакция сопровождается внутримолекулярными окислительно-восстановительными процессами, в результате которых образуется фосфоенолпируват (фосфоенолпировиноградная кислота) с высокоэнергетической связью во втором атоме углерода и отщепляется молекула воды

фермент: енолаза
10. Реакция субстратного фосфорилирования
В ходе десятой реакции фосфорильная группа переносится на ADP. При этом синтезируется АТР и пируват (пировиноградная кислота). Эта реакция также необратима, поскольку высокоэкзергонична.
фермент: пируваткиназа
11. Реакция обратимого восстановления пировиноградной кислоты до молочной кислоты (в анаэробных условиях)
Если после гликолиза следует аэробное расщепление, пируват мигрирует в матрикс митохондрий, где, взаимодействуя с коэнзимом-А, участвует в образовании ацетил-СоА. В анаэробных условиях пируват при участии NADH восстанавливается до лактата (молочной кислоты), который при этом является конечным продуктом гликолиза. Затем в аэробных условиях лактат может обратно превратиться в пируват и окислиться в митохондриях.

фермент: лактатдегидрогеназа
1. Биология для поступающих в вузы / Г.Л. Билич, В.А. Крыжановский. — 2008.
2. Биология в таблицах и схемах / Спб. — 2004.
3. Биохимия в схемах и таблицах / И. В. Семак – Минск — 2011.
Источник
Универсальным источником энергии во всех клетках служит АТФ (аденозинтрифосфат, или аденозинтрифосфорная кислота).
Все энергетические затраты любой клетки обеспечиваются за счёт универсального энергетического вещества — АТФ.
АТФ синтезируется в результате реакции фосфорилирования, то есть присоединения одного остатка фосфорной кислоты к молекуле АДФ (аденозиндифосфата):
АДФ + H3PO4+ 40 кДж = АТФ + H2O.
Энергия запасается в форме энергии химических связей АТФ. Химические связи АТФ, при разрыве которых выделяется много энергии, называются макроэргическими.
При распаде АТФ до АДФ клетка за счёт разрыва макроэргической связи получит приблизительно (40) кДж энергии.
Энергия для синтеза АТФ из АДФ выделяется в процессе диссимиляции.
Энергетический обмен (диссимиляция, катаболизм) — это совокупность химических реакций постепенного распада органических соединений, сопровождающихся высвобождением энергии, часть которой расходуется на синтез АТФ.
В зависимости от среды обитания организма, диссимиляция может проходить в два или в три этапа.
Процессы расщепления органических соединений у аэробных организмов происходят в три этапа: подготовительный, бескислородный и кислородный.
В результате этого органические вещества распадаются до простейших неорганических соединений.
У анаэробных организмов, обитающих в бескислородной среде и не нуждающихся в кислороде (а также у аэробных организмов при недостатке кислорода), диссимиляция происходит в два этапа: подготовительный и бескислородный.
В двухэтапном энергетическом обмене энергии запасается гораздо меньше, чем в трёхэтапном.
Первый этап — подготовительный
Подготовительный этап заключается в распаде крупных органических молекул до более простых: полисахаридов — до моносахаридов, липидов — до глицерина и жирных кислот, белков — до аминокислот.
Этот процесс называется пищеварением. У многоклеточных организмов он осуществляется в желудочно-кишечном тракте с помощью пищеварительных ферментов. У одноклеточных организмов — происходит под действием ферментов лизосом.
В ходе биохимических реакций, происходящих на этом этапе, энергии выделяется мало, она рассеивается в виде тепла, и АТФ не образуется.
Второй этап — бескислородный (гликолиз)
Второй (бескислородный) этап заключается в ферментативном расщеплении органических веществ, которые были получены в ходе подготовительного этапа. Кислород в реакциях этого этапа не участвует.
Биологический смысл второго этапа заключается в начале постепенного расщепления и окисления глюкозы с накоплением энергии в виде (2) молекул АТФ.
Процесс бескислородного расщепления глюкозы называется гликолиз.
Гликолиз происходит в цитоплазме клеток.
Он состоит из нескольких последовательных реакций превращения молекулы глюкозы C6H12O6 в две молекулы пировиноградной кислоты — ПВК C3H4O3 и две молекулы АТФ (в виде которой запасается примерно (40) % энергии, выделившейся при гликолизе). Остальная энергия (около (60) %) рассеивается в виде тепла.
C6H12O6+2H3PO4+2АДФ=2C3H4O3+2АТФ +2H2O.
Получившаяся пировиноградная кислота при недостатке кислорода в клетках животных, а также клетках многих грибов и микроорганизмов, превращается в молочную кислоту C3H6O3.
HOOC−CO−CH3пировиноградная кислота→НАД⋅H+H+лактатдегидрогеназаHOOC−CHOH−CH3молочная кислота.
В мышцах человека при больших нагрузках и нехватке кислорода образуется молочная кислота и появляется боль. У нетренированных людей это происходит быстрее, чем у людей тренированных.
При недостатке кислорода в клетках растений, а также в клетках некоторых грибов (например, дрожжей), вместо гликолиза происходит спиртовое брожение: пировиноградная кислота распадается на этиловый спирт C2H5OH и углекислый газ CO2:
C6H12O6+2H3PO4+2АДФ=2C2H5OH+2CO2+2АТФ+2H2O.
Третий этап — кислородный
В результате гликолиза глюкоза распадается не до конечных продуктов (CO2 и H2O), а до богатых энергией соединений (молочная кислота, этиловый спирт) которые, окисляясь дальше, могут дать её в больших количествах. Поэтому у аэробных организмов после гликолиза (или спиртового брожения) следует третий, завершающий этап энергетического обмена — полное кислородное расщепление, или клеточное дыхание.
Этот этап происходит на кристах митохондрий.
Третий этап, так же как и гликолиз, является многостадийным и состоит из двух последовательных процессов — цикла Кребса и окислительного фосфорилирования.
Третий (кислородный) этап заключается в том, что при кислородном дыхании ПВК окисляется до окончательных продуктов — углекислого газа и воды, а энергия, выделяющаяся при окислении, запасается в виде (36) молекул АТФ ((2) молекулы в цикле Кребса и (34) молекулы в ходе окислительного фосфорилирования).
Этот этап можно представить себе в следующем виде:
2C3H4O3+6O2+36H3PO4+36АДФ=6CO2+42H2O+36АТФ.
Вспомним, что ещё две молекулы АТФ запасаются в ходе бескислородного расщепления каждой молекулы глюкозы (на втором, бескислородном, этапе). Таким образом, в результате полного расщепления одной молекулы глюкозы образуется (38) молекул АТФ.
Суммарная реакция энергетического обмена:
C6H12O6+6O2=6CO2+6H2O+38АТФ.
Для получения энергии в клетках, кроме глюкозы, могут быть использованы и другие вещества: липиды, белки. Однако ведущая роль в энергетическом обмене у большинства организмов принадлежит сахарам.
Источники:
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. 9 класс // ДРОФА.
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. Общая биология (базовый уровень) 10–11 класс // ДРОФА.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
Источник
Энергетический обмен
Энергетический обмен (катаболизм, диссимиляция) — совокупность реакций расщепления органических веществ, сопровождающихся выделением энергии. Энергия, освобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме АТФ и других высокоэнергетических соединений. АТФ — универсальный источник энергообеспечения клетки. Синтез АТФ происходит в клетках всех организмов в процессе фосфорилирования — присоединения неорганического фосфата к АДФ.
У аэробных организмов (живущих в кислородной среде) выделяют три этапа энергетического обмена: подготовительный, бескислородное окисление и кислородное окисление; у анаэробных организмов (живущих в бескислородной среде) и аэробных при недостатке кислорода — два этапа: подготовительный, бескислородное окисление.
Подготовительный этап
Заключается в ферментативном расщеплении сложных органических веществ до простых: белковые молекулы — до аминокислот, жиры — до глицерина и карбоновых кислот, углеводы — до глюкозы, нуклеиновые кислоты — до нуклеотидов. Распад высокомолекулярных органических соединений осуществляется или ферментами желудочно-кишечного тракта или ферментами лизосом. Вся высвобождающаяся при этом энергия рассеивается в виде тепла. Образовавшиеся небольшие органические молекулы могут быть использованы в качестве «строительного материала» или могут подвергаться дальнейшему расщеплению.
Бескислородное окисление, или гликолиз
Этот этап заключается в дальнейшем расщеплении органических веществ, образовавшихся во время подготовительного этапа, происходит в цитоплазме клетки и в присутствии кислорода не нуждается. Главным источником энергии в клетке является глюкоза. Процесс бескислородного неполного расщепления глюкозы — гликолиз.
Потеря электронов называется окислением, приобретение — восстановлением, при этом донор электронов окисляется, акцептор восстанавливается.
Следует отметить, что биологическое окисление в клетках может происходить как с участием кислорода:
А + О2 → АО2,
так и без его участия, за счет переноса атомов водорода от одного вещества к другому. Например, вещество «А» окисляется за счет вещества «В»:
АН2 + В → А + ВН2
или за счет переноса электронов, например, двухвалентное железо окисляется до трехвалентного:
Fe2+ → Fe3+ + e—.
Гликолиз — сложный многоступенчатый процесс, включающий в себя десять реакций. Во время этого процесса происходит дегидрирование глюкозы, акцептором водорода служит кофермент НАД+ (никотинамидадениндинуклеотид). Глюкоза в результате цепочки ферментативных реакций превращается в две молекулы пировиноградной кислоты (ПВК), при этом суммарно образуются 2 молекулы АТФ и восстановленная форма переносчика водорода НАД·Н2:
С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ → 2С3Н4О3 + 2АТФ + 2Н2О + 2НАД·Н2.
Дальнейшая судьба ПВК зависит от присутствия кислорода в клетке. Если кислорода нет, у дрожжей и растений происходит спиртовое брожение, при котором сначала происходит образование уксусного альдегида, а затем этилового спирта:
- С3Н4О3 → СО2 + СН3СОН,
- СН3СОН + НАД·Н2 → С2Н5ОН + НАД+.
У животных и некоторых бактерий при недостатке кислорода происходит молочнокислое брожение с образованием молочной кислоты:
С3Н4О3 + НАД·Н2 → С3Н6О3 + НАД+.
В результате гликолиза одной молекулы глюкозы высвобождается 200 кДж, из которых 120 кДж рассеивается в виде тепла, а 80% запасается в связях АТФ.

Кислородное окисление, или дыхание
Заключается в полном расщеплении пировиноградной кислоты, происходит в митохондриях и при обязательном присутствии кислорода.
Пировиноградная кислота транспортируется в митохондрии (строение и функции митохондрий — лекция №7). Здесь происходит дегидрирование (отщепление водорода) и декарбоксилирование (отщепление углекислого газа) ПВК с образованием двухуглеродной ацетильной группы, которая вступает в цикл реакций, получивших название реакций цикла Кребса. Идет дальнейшее окисление, связанное с дегидрированием и декарбоксилированием. В результате на каждую разрушенную молекулу ПВК из митохондрии удаляется три молекулы СО2; образуется пять пар атомов водорода, связанных с переносчиками (4НАД·Н2, ФАД·Н2), а также одна молекула АТФ.

Суммарная реакция гликолиза и разрушения ПВК в митохондриях до водорода и углекислого газа выглядит следующим образом:
С6Н12О6 + 6Н2О → 6СО2 + 4АТФ + 12Н2.
Две молекулы АТФ образуются в результате гликолиза, две — в цикле Кребса; две пары атомов водорода (2НАДЧН2) образовались в результате гликолиза, десять пар — в цикле Кребса.
Последним этапом является окисление пар атомов водорода с участием кислорода до воды с одновременным фосфорилированием АДФ до АТФ. Водород передается трем большим ферментным комплексам (флавопротеины, коферменты Q, цитохромы) дыхательной цепи, расположенным во внутренней мембране митохондрий. У водорода отбираются электроны, которые в матриксе митохондрий в конечном итоге соединяются с кислородом:
О2 + e— → О2—.
Купить проверочные работы
и тесты по биологии




Протоны закачиваются в межмембранное пространство митохондрий, в «протонный резервуар». Внутренняя мембрана непроницаема для ионов водорода, с одной стороны она заряжается отрицательно (за счет О2—), с другой — положительно (за счет Н+). Когда разность потенциалов на внутренней мембране достигает 200 мВ, протоны проходят через канал фермента АТФ-синтетазы, образуется АТФ, а цитохромоксидаза катализирует восстановление кислорода до воды. Так в результате окисления двенадцати пар атомов водорода образуется 34 молекулы АТФ.

1 — наружная мембрана; 2 — межмембранное пространство, протонный резервуар;
3 — цитохромы; 4 — АТФ-синтетаза.
При перфорации внутренних митохондриальных мембран окисление НАД·Н2 продолжается, но АТФ-синтетаза не работает и образования АТФ в дыхательной цепи не происходит, энергия рассеивается в форме тепла (клетки «бурого жира» млекопитающих).
Суммарная реакция расщепления глюкозы до углекислого газа и воды выглядит следующим образом:
С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 38АТФ + Qт,
где Qт — тепловая энергия.
Перейти к лекции №10 «Понятие об обмене веществ. Биосинтез белков»
Перейти к лекции №12 «Фотосинтез. Хемосинтез»
Смотреть оглавление (лекции №1-25)
Источник