
Какие свойствами обладает цитоплазматическая

Наследственность цитоплазматическая (греч. kytos вместилище, клетка + plasma вылепленное, оформленное; син.: неменделевская наследственность, наследственность плазматическая, внехромосомная наследственность, нехромосомная наследственность, внеядерная наследственность) — передача потомству отдельных признаков и свойств, обусловленных нехромосомными (цитоплазматическими) преемственными структурными элементами клетки. Этот процесс совершается частично автономно, частично — в зависимости от клеточного ядра.
Наследственность цитоплазматическая, в отличие от менделевской наследственности (см. Менделя законы), характеризуется тем, что цитоплазматические наследственные факторы — цитоплазматические гены, или плазмогены, составляют плазмон и не расщепляются, а передаются всему потомству однородительским путем. Т. к. яйцеклетка содержит цитоплазмы во много раз больше, чем спермий, то при слиянии обеих гамет женская гамета вносит гораздо больший вклад в цитоплазму зиготы, чем мужская. Поэтому практически все цитоплазматические гены передаются потомству по материнской линии.
К преемственным структурным элементам цитоплазмы, являющимся носителями плазмогенов, относят митохондрии (см.) всех эукариотов, т. е. организмов, клетки к-рых в противоположность прокариотам содержат типичное ядро, окруженное мембраной, а также хлоропласты зеленых растений и водорослей, кинетопласт жгутиковых простейших (трипаносомид) и другие органоиды цитоплазмы. У бактерий и нек-рых низших эукариотов к этим же преемственным структурным элементам условно относят эписомы (см.) и плазмиды (см.). Наконец, к ним же могут быть условно отнесены и паразиты (эндосибионты), обитающие в клетках эукариотов.
Первыми изученными примерами такого сожительства могут служить частички Каппа — разновидность бактерий, утративших способность к самостоятельной жизни и поселившихся в теле парамеций, куда они выделяют особый токсин, убивающий особей, лишенных этих частиц. У дрозофилы обнаружен другой такой эндосимбионт—вирус Сигма, сообщающий плодовым мушкам аномальную чувствительность к углекислому газу. Эти признаки передаются потомству только в том случае, если цитоплазма зиготы получена от зараженного родителя.
Явление Наследственности цитоплазматической описано Корренсом (К. Correns) и Бауром (Е. Ваш) в 1908 г. Они установили, что у цветковых растений наследование признака пестролистности и передача пластид («пластидная наследственность») осуществляется гл. обр. или даже полностью через женские репродуктивные клетки, т. е. идет по однородительской, а не по менделевской схеме. Впоследствии подобные явления были обнаружены у других высших растений, водорослей, грибов, простейших, насекомых, у многоклеточных животных, в т. ч. и у высших млекопитающих и человека.
Цитоплазматическая природа наследственности доказывается различными способами, чаще всего — методами реципрокных и повторных обратных скрещиваний (см. Скрещивание). Многократное повторение скрещиваний типа А♀ x B♂ x B♂ x B♂ в конце концов приведет к тому, что материнские ядра вида А у гибридов будут полностью заменены на отцовские ядра вида В, а цитоплазма вида А сохранится. Различия, обнаруженные в потомстве от этих скрещиваний, несомненно, будут указывать на цитоплазматическую природу наследования. Этот метод позволяет отличить истинное плазматическое наследование от так наз. материнского эффекта, к-рый по нек-рым проявлениям напоминает генетическое действие цитоплазмы, но на самом деле объясняется предетерминацией цитоплазмы яйцеклетки генотипом материнского организма до оплодотворения.
Доказательством того, что цитоплазматические структуры, плазмогены, определяют развитие нек-рых признаков организма, может служить следующее: обнаружение разнообразных мутаций, передающихся только через цитоплазму; открытие в составе субклеточных органелл специфических ДНК, рибосомных, транспортных и матричных РНК и особого аппарата синтеза белка; установление непосредственной связи между выпадением или изменением полидзоксирибонуклеотидных последовательностей в молекуле ДНК органелл и изменением фенотипа у цитоплазматических мутантов; обнаружение трансмиссии, сегрегации и рекомбинации плазмогенов.
Мутации, изменяющие или полностью нарушающие функции и характерные свойства митохондрий (транспорт электронов в дыхательной цепи, окцелительное фосфорилирование, чувствительность к нек-рым ядам и др.), были выделены у грибов, простейших и у многоклеточных животных. Особый интерес представляют мутанты, устойчивые к антибактериальным антибиотикам, у низших эукариотов и в культурах клеток млекопитающих. Существование таких мутантов указывает на то, что норма реакции клеток состоит в чувствительности к этим агентам и что токсичность антибиотиков для человека объясняется именно повреждением митохондрий. Набор мутаций. изменяющих частные реакции в обмене веществ и биосинтетической активности митохондрий, дает возможность расчленить их функции на отдельные этапы и таким путем подойти к анализу этих функций.
Генетический анализ позволил выявить у дрожжей Saccharomyces cerevisiae сегрегацию и рекомбинацию митохондриальных генов. Это сделало возможным установление основных особенностей поведения митохондриальных генов, отличающих их от поведения ядерных генов: наличие сегрегации в ходе митоза (см.); отсутствие сегрегации при мейозе (см.) и полярность рекомбинаций, т. е. преимущественная передача потомству генотипа митохондрий одного из родителей (половые различия у митохондрий). На основе анализа делеций (см.) и точковых мутаций митохондрий дрожжевых клеток было произведено картирование значительного числа локусов митохондриального генома.
Митохондрии не образуются de novo, а размножаются путем деления клеток. Рекомбинация митохондриальных генов, по крайней мере у дрожжей, становится возможной благодаря «слиянию» митохондрий, диссоциации их мембран и наследственному контакту между молекулами митохондриальных ДНК.
Особое значение (в т. ч. и для человека) имеют исследования, выполненные на высших животных. Так, сегрегация и рекомбинация митохондриальных генов установлены в культуре гибридных соматических клеток «человек-мышь». Произведено картирование генов, определяющих рибосомные и транспортные РНК в культуре клеток HeLa и Xenopus laevis с применением молекулярной гибридизации РНК — ДНК, электронной микроскопии и рестрикционного анализа. У млекопитающих продемонстрирована внутривидовая гетерогенность митохондриальной ДНК и установлено, что специфичность этой ДНК передается в поколениях не через ядра половых клеток, а через цитоплазму.
Результаты исследований генетических функций цитоплазмы не поколебали основного принципа генетики, подавляющее большинство генетической информации несут хромосомные гены клеточного ядра, заключенные в ядерной ДНК (см. Дезоксирибонуклеиновые кислоты). Эти исследования только показали, что часть генетической информации содержится в ДНК цитоплазмы, в т. ч. в митохондриальной ДНК (мтДНК); мтДНК представлена кольцевыми ковалентно-замкнутыми молекулами длиной от 5 мкм у высших животных до 25 мкм у дрожжей. Молекулы мтДНК образуются только из мтДНК, точно копируя последовательность их полидезоксирибонуклео-тидов, и не дублируют последовательность нуклеотидов ядерной ДНК. Гибридизационные исследования демонстрируют отсутствие гомологии между мтДНК и ядерной ДНК.
Размер молекул митохондриальной ДНК невелик, и геном митохондрий не может полностью обеспечить процесс воспроизведения структуры митохондрий. Мол. вес (масса) митохондриальной ДНК колеблется от 107 у животных до 5 * 107 у дрожжей. Кодирующая емкость митохондриальной ДНК определяет структуру только 8—15% структурных компонентов митохондрий. К их числу относятся большая и малая рибосомные РНК, около двадцати транспортных РНК и 8—12 матричных РНК. Митохондриальные матричные РНК, в свою очередь, кодируют ряд компонентов внутренних мембран: три субъединицы цитохромоксидазы (КФ 1.9.3.1) из семи, одну субъединицу цитохрома b из двух, четыре субъединицы АТФ-синтетазы (КФ 3.6.1.3) из десяти и еще неидентифицированные продукты, составляющие всего ок. 40% кодирующей емкости митохондриальной ДНК. К числу таких неидентифицированных продуктов относятся и вещества, контролирующие рост и другие функции клетки как целого (экстрамитохондриальные функции митохондрий). Продукты митохондриального генома синтезируются митохондриальными рибосомами (см.), напоминающими по ряду свойств бактериальные рибосомы. Однако преобладающая часть полипептидов интегральных компонентов митохондрий кодируется хромосомными генами и синтезируется в рибосомах цитоплазмы. Эти ядерно-цитоплазматические продукты доставляются в митохондрии и здесь соединяются с полипептидам и, синтезированными в митохондриальных рибосомах, в результате чего образуются единые олигомерные ферментные комплексы. Зависимость митохондрий от ядра этим не ограничивается. Другим немаловажным фактором является контроль ядра над процессами репликации, рекомбинации, транскрипции и трансляции митохондриальной ДНК. Контроль ядра заключается в том, что ферменты, участвующие в этих процессах, являются продуктами ядерных генов. Все это свидетельствует о том, что генетические функции митохондрий только отчасти осуществляются автономно, в той мере, в какой это определяется специфичностью митохондриальной ДНК. Но и митохондрии, в свою очередь, вносят некоторый вклад в функции ядерно-цитоплазматической системы.
Об этом свидетельствуют митохондриальные мутации, нарушающие ядерную систему у дрожжей и парамеций. Т. о., ядро и митохондрии образуют интегрированную генетическую систему клетки, причем вклад митохондрий, хотя относительно и очень мал, но уникален и не может компенсироваться активностью ядра.
Генетическая система хлоропластов, определяющая функции преобразования солнечной энергии в энергию хим. реакций в зеленом покрове Земли, в принципе организована подобно митохондриальной системе. Двойной генетический контроль над функциями клетки с участием ядра и цитоплазмы обеспечивает надежность и точность регулирования процессов превращения энергии в органическом мире.
См. также Изменчивость, Наследование, Наследственность.
Библиография: Гаузе Г. Г. Митохондриальная ДНК, М., 1978; Генетические функции органоидов цитоплазмы, под ред. С. А. Нейфаха, Д., 1974; Каллиникова В. Д. Клеточная органелла кинетопласт, Л., 1977; Молекулярная генетика митохондрий, под ред. С. А. Нейфаха и А. С. Трошина, Л., 1977; Beale G. а. Knowles J. Extranuclear genetics, L., 1978; Borst P. a. Grivell L. A. The mitochondrial genome of yeast, Cell, v. 15, p. 705, 1978; The genetic function of mitochondrial DNA, ed. by G. Saccone a. A. M. Kroon, p. 15, Amsterdam, 1976; Whitehouse H. L. K. Towards an understanding of the mechanism of heredity, L., 1969.
С. А. Нейфах.
Источник
Наружная цитоплазматическая мембрана представляет собой тончайшую пленку. Ее толщина – порядка 7-10 нм. Просматривается пленка только в электронный микроскоп.
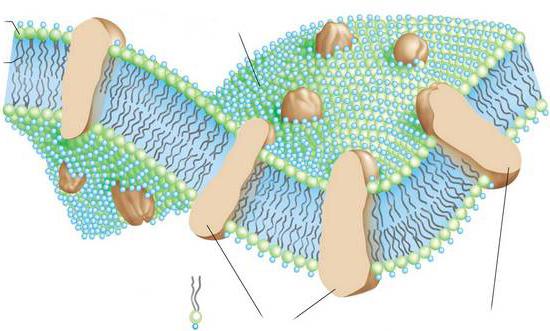
Далее рассмотрим, что собой представляет цитоплазматическая мембрана. Функции пленки также будут описаны в статье.
Структура
Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные (“боящиеся влаги”) и гидрофильные (“любящие воду”) концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой.
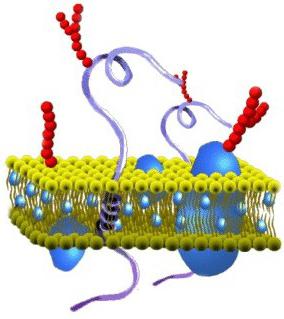
Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды – основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Цитоплазматическая мембрана: функции
Основными задачами являются:
- Барьерная. Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
- Транспортная. Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи
- Матричная. Эта функция обеспечивает определенную ориентацию и взаиморасположение белков мембраны, а также оптимальное их взаимодействие.
- Механическая. За счет нее обеспечивается автономность клетки, внутренних структур. Также осуществляется соединение элемента с прочими аналогичными.
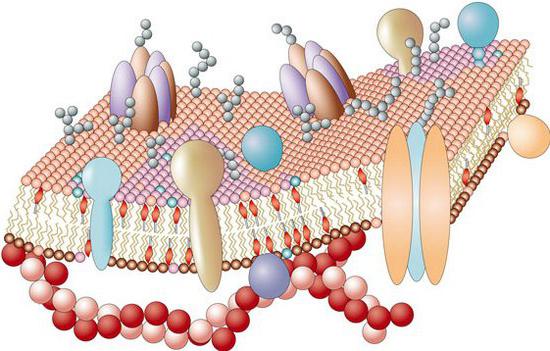
- Энергетическая. На фоне фотосинтеза в хлоропластах и при осуществлении клеточного дыхания в мембранах активны системы энергетического переноса. В них также участвуют и белковые соединения.
- Рецепторная. Ряд белков, которые присутствуют в мембране, обеспечивает восприятие различных сигналов. К примеру, циркулирующие в крови стероиды оказывают воздействие только на те клетки-мишени, которые обладают соответствующими гормонам рецепторами. Химические соединения, обеспечивающие проведение импульсов (нейромедиаторы), также связываются с помощью особых белков клеток-мишеней.
Особые свойства
К специфическим функциям мембраны относят:
- Ферментативную. Зачастую белки, которые содержит цитоплазматическая мембрана, выступают в качестве ферментов.
- Генерацию и проведение биопотенциалов.
- Маркировку. Цитоплазматическая мембрана включает в свой состав особые антигены. Они действуют как маркеры-“ярлыки”. Благодаря им осуществляется распознание клеток. Маркеры представляют собой гликопротеины – белки, содержащие разветвленные олигосахаридные боковые цепи. Они выступают в качестве “антенн”.
Благодаря огромному количеству вариантов боковых цепей для того или другого типа клеток может быть сформирован особый маркер. При их помощи распознанные друг другом элементы начинают действовать согласованно. К примеру, так происходит при образовании тканей и органов. Маркировка также позволяет иммунитету определить чужеродные антигены.
Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза.
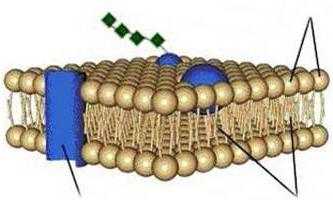
В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Модели
Их существует несколько:
- “Бутербродная модель”. Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- “Жидкостно-мозаичная структура”. Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- “Белково-кристаллическая структура”. В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.
Источник
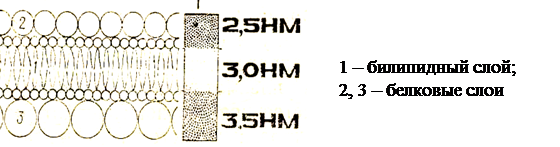
Цитоплазматическая мембрана или плазмалемма (лат. membrana – кожица, плёнка) – тончайшая пленка (7–10нм), отграничивающая внутреннее содержимое клетки от окружающей среды, видна только в электронный микроскоп.
По химической организации плазмалемма представляет липопротеидный комплекс – молекулы липидов и белков.
Её основу составляет липидный бислой, состоящий из фосфолипидов, кроме этого, в мембранах присутствуют гликолипиды и холестерол. Все они обладают свойством амфипатричности, т.е. у них есть гидрофильные («любящие воду») и гидрофобные («боящиеся воды») концы. Гидрофильные полярные «головки» липидных молекул (фосфатная группа) обращены кнаружи мембраны, а гидрофобные неполярные «хвосты» (остатки жирных кислот) – друг к другу, что создает биполярный липидный слой. Молекулы липидов подвижны и могут перемещаться в своем монослое или редко – из одного монослоя в другой. Монослои липидов обладают ассиметричностью, т. е. отличаются по составу липидов, что придает специфичность мембранам даже в пределах одной клетки. Бислой липидов может находиться в состоянии жидкого или твердого кристалла.
Вторым обязательным компонентом плазмалеммы являются белки. Многие мембранные белки способны перемещаться в плоскости мембраны или вращаться вокруг своей оси, но не могут переходить с одной стороны бислоя липидов на другой.
Липиды обеспечивают основные структурные особенности мембраны, а белки – её функции. Функции мембранных белков различны: поддержание структуры мембран, получение и преобразование сигналов из окружающей среды, транспорт некоторых веществ, катализ реакций, происходящих на мембранах.
Различают несколько моделей строения цитоплазматической мембраны.
①. БУТЕРБРОДНАЯ МОДЕЛЬ(белки – липиды – белки)
В 1935г. английские ученые Даниэли и Даусон высказали идею о послойном расположении в мембранемолекул белков (темные слои в электронном микроскопе), которые залегают снаружи, и молекул липидов (светлый слой) – внутри. Длительное время существовало представление о едином трехслойном строении всех биологических мембран.
При детальном изучении мембраны с помощью электронного микроскопа оказалось, что светлый слой на самом деле представлен двумя слоями фосфолипидов – это билипидный слой, причем водорастворимые его участки – гидрофильные головки направлены к белковому слою, а нерастворимые (остатки жирных кислот) – гидрофобные хвосты обращены друг к другу.
 |
Однако уже с середины 60-х годов начали накапливаться факты против унитарной «бутербродной» модели. В частности, по одним данным, не все мембраны имели четкую трехслойную структуру при электронно-микроскопическом исследовании; по другим – значительная часть мембранных белков имела глобулярную структуру, а не ламеллярную, как в постулируемой модели. Наконец, среди многочисленных моделей мембран, предложенных в середине 60-х годов, начали выделяться те, в которых доказывалось наличие гидрофобно-гидрофильных взаимодействий не только между липидными молекулами, но и между липидами и белками.
 ②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
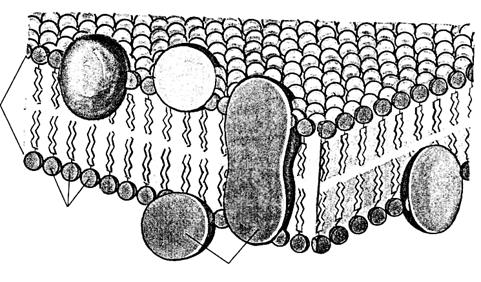
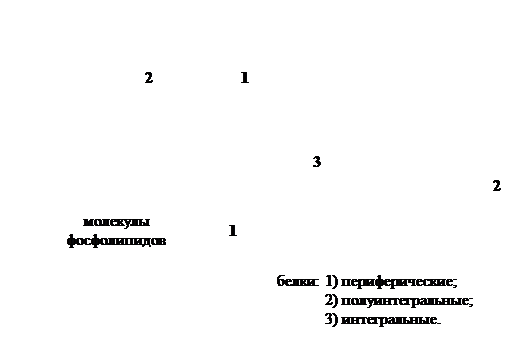
В 1972г.Сингер и Николсонописали модель мембраны, которая получила широкое признание. Согласно этой модели молекулы белков не образуют сплошного слоя, а погружены в биполярный липидный слой на разную глубину в виде мозаики. Глобулы белковых молекул, подобно айсбергам, погружены в «океан»
липидов: одни находятся на поверхности билипидного слоя – периферические белки, другие погружаются в него наполовину – полуинтегральные белки, третьи – интегральные белки – пронизывают его насквозь, формируя гидрофильные поры. Периферические белки, находясь на поверхности билипидного слоя, связаны с головками липидных молекул электростатическими взаимодействиями. Но они никогда не образуют сплошного слоя и, по сути дела, не являются белками собственно мембраны, а, скорее, связывают ее с надмембранной или субмембранной системой поверхностного аппарата клетки.
Основную роль в организации собственно мембраны играют интегральные и полуинтегральные (трансмембранные) белки, имеющие глобулярную структуру и связанные с липидной фазой гидрофильно-гидрофобными взаимодействиями. Молекулы белков, как и липиды, обладают амфипатричностью и своими гидрофобными участками взаимодействуют с гидрофобными хвостами билипидного слоя, а гидрофильные участки обращены к водной среде и образуют с водой водородные связи.
③. БЕЛКОВО-КРИСТАЛЛИЧЕСКАЯ МОДЕЛЬ(модель липопротеинового коврика)
Мембраны образованы переплетением липидных и белковых молекул, объединяющихся между собой на основе гидрофильно-
гидрофобных взаимодействий.
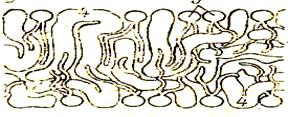
Белковые молекулы, как штифты, пронизывают слой липидов и выполняют в составе мембраны функцию каркаса. После обработки мембраны жирорастворимыми веществами белковый каркас сохраняется, что доказывает взаимосвязь между молекулами белков в мембране. По-видимому, эта модель реализуется лишь в отдельных специальных участках некоторых мембран, где требуется жесткая структура и тесные стабильные взаимоотношения между липидами и белками (например, в области расположения фермента Na-К –АТФ-азы).
Самой универсальной моделью, отвечающей термодинамическим принципам (принципам гидрофильно-гидрофобных взаимодействий), морфо-биохимическим и экспериментально-цитологическим данным, является жидкостно-мозаичная модель. Однако все три модели мембран не исключают друг друга и могут встречаться в разных участках одной и той же мембраны в зависимости от функциональных особенностей данного участка.
СВОЙСТВА МЕМБРАНЫ
1. Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою структуру, т.к. молекулы липидов на основе своих физико-химических свойств собираются в биполярный слой, в который затем встраиваются молекулы белков.
2. Текучесть. Мембрана не является жесткой структурой, большая часть входящих в её состав белков и липидов может перемещаться в плоскости мембраны, они постоянно флюктуируют за счет вращательных и колебательных движений. Это определяет большую скорость протекания химических реакций на мембране.
3. Полупроницаемость. Мембраны живых клеток пропускают, помимо воды, лишь определённые молекулы и ионы растворённых веществ. Это обеспечивает поддержание ионного и молекулярного состава клетки.
4. Мембрана не имеет свободных концов. Она всегда замыкается в пузырьки.
5. Асимметричность. Состав наружного и внутреннего слоев как белков, так и липидов различен.
6. Полярность. Внешняя сторона мембраны несёт положительный заряд, а внутренняя – отрицательный.
ФУНКЦИИ МЕМБРАНЫ
1) Барьерная –плазмалемма отграничивает цитоплазму и ядро от внешней среды. Кроме того, мембрана делит внутреннее содержимое клетки на отсеки (компартменты), в которых зачастую протекают противоположные биохимические реакции.
2) Рецепторная(сигнальная) – благодаря важному свойству белковых молекул – денатурации, мембрана способна улавливать различные изменения в окружающей среде. Так, при воздействии на мембрану клетки различных средовых факторов (физических, химических, биологических) белки, входящие в ее состав, меняют свою пространственную конфигурацию, что служит своеобразным сигналом для клетки. Это обеспечивает связь с внешней средой, распознавание клеток и их ориентацию при формировании тканей и т.д. С этой функцией связана деятельность различных регуляторных систем и формирование иммунного ответа.
3) Обменная – в состав мембраны входят не только структурные белки, которые образуют ее, но и ферментативные, являющиеся биологическими катализаторами. Они располагаются на мембране в виде «каталитического конвейера» и определяют интенсивность и направленность реакций метаболизма.
4) Транспортная – молекулы веществ, диаметр которых не превышает 50 нм, могут проникать путем пассивного и активного транспорта через поры в структуре мембраны. Крупные вещества попадают в клетку путем эндоцитоза (транспорт в мембранной упаковке), требующего затраты энергии. Его разновидностями являются фаго- и пиноцитоз.
Пассивный транспорт – вид транспорта, в котором перенос веществ осуществляется по градиенту химической или электрохимической концентрации без затраты энергии АТФ. Выделяют два вида пассивного транспорта: простая и облегченная диффузия. Диффузия – это перенос ионов или молекул из зоны более высокой их концентрации в зону более низкой концентрации, т.е. по градиенту.
Простая диффузия – ионы солей и вода проникают через трансмембранные белки или жирорастворимые вещества по градиенту концентрации.
Облегченная диффузия – специфические белки-переносчики связывают вещество и переносят его через мембрану по принципу «пинг-понга». Таким способом через мембрану проходят сахара и аминокислоты. Скорость такого транспорта значительно выше, чем простой диффузии. Кроме белков- переносчиков, в облегченной диффузии принимают участие некоторые антибиотики – например, грамитидин и ваномицин. Поскольку они обеспечивают транспорт ионов, их называют ионофорами.
Активный транспорт – это вид транспорта, при котором расходуется энергия АТФ, он идёт против градиента концентрации. В нем принимают участие ферменты АТФ-азы. В наружной клеточной мембране находятся АТФ-азы, которые осуществляют перенос ионов против градиента концентрации, это явление называется ионным насосом. Примером является натрий-калиевый насос. В норме в клетке больше ионов калия, во внешней среде – ионов натрия. Поэтому по законам простой диффузии калий стремится из клетки, а натрий – в клетку. В противовес этому натрий-калиевый насос накачивает против градиента концентрации в клетку ионы калия, а ионы натрия выносит во внешнюю среду. Это позволяет поддерживать постоянство ионного состава в клетке и её жизнеспособность. В животной клетке одна треть АТФ расходуется на работу натрий-калиевого насоса.
Разновидностью активного транспорта является транспорт в мембранной упаковке – эндоцитоз. Крупные молекулы биополимеров не могут проникать через мембрану, они поступают в клетку в мембранной упаковке. Различают фагоцитоз и пиноцитоз. Фагоцитоз – захват клеткой твердых частиц, пиноцитоз – жидких частиц. В этих процессах выделяют стадии:
1) узнавание рецепторами мембраны вещества; 2) впячивание (инвагинация) мембраны с образованием везикулы (пузырька); 3) отрыв пузырька от мембраны, слияние его с первичной лизосомой и восстановление целостности мембраны; 4) выделение непереваренного материала из клетки (экзоцитоз).
Эндоцитоз является способом питания для простейших. У млекопитающих и человека имеется ретикуло-гистио-эндотелиальная система клеток, способная к эндоцитозу – это лейкоциты, макрофаги, клетки Купфера в печени.
ОСМОТИЧЕСКИЕ СВОЙСТВА КЛЕТКИ
Осмос – односторонний процесс проникновения воды через полупроницаемую мембрану из области с меньшей концентрацией раствора в область с более высокой концентрацией. Осмос обусловливает осмотическое давление.
Диализ – односторонняя диффузия растворенных веществ.
Раствор, в котором осмотическое давление такое же, как и в клетках, называют изотоническим. При погружении клетки в изотонический раствор её объем не изменяется. Изотонический раствор называют физиологическим – это 0,9% раствор хлорида натрия, который широко применяется в медицине при сильном обезвоживании и потери плазмы крови.
Раствор, осмотическое давление которого выше, чем в клетках, называют гипертоническим. Клетки в гипертоническом растворе теряют воду и сморщиваются. Гипертонические растворы широко применяются в медицине. Марлевая повязка, смоченная в гипертоническом растворе, хорошо впитывает гной.
Раствор, где концентрация солей ниже, чем в клетке, называют гипотоническим. При погружении клетки в такой раствор вода устремляется в нее. Клетка набухает, ее тургор увеличивается, и она может разрушиться. Гемолиз – разрушение клеток крови в гипотоническом растворе.
Осмотическое давление в организме человека в целом регулируется системой органов выделения.
Источник