
Каким свойством не обладают нервные центры

Учение о рефлекторной деятельности ЦНС привело к развитию представления о нервном центре.
Нервным центром называют совокупность нейронов, необходимых для осуществления определенного рефлекса или регуляции той или иной функции.
Не следует понимать нервный центр как что-то узко локализованное в одном участке ЦНС. Понятие анатомическое по отношению к нервному центру рефлекса неприменимо потому, что в осуществлении любого сложного рефлекторного акта принимает участие всегда целая констелляция нейронов, расположенных на разных этажах нервной системы. Опыты с раздражением или перерезкой ЦНС показывают лишь, что отдельные нервные образования обязательны для осуществления того или иного рефлекса, а другие необязательны, хотя и участвуют при обычных условиях в рефлекторной деятельности. Примером служит дыхательный центр, в который в настоящее время включают не только “центр дыхания” продолговатого мозга, но и пневмотаксический центр моста, нейроны ретикулярной формации, коры и мотонейроны дыхательных мышц.
Нервные центры обладают рядом характерных свойств, определяемых свойствами составляющих его нейронов, особенностями синаптической передачи нервных импульсов и структурой нейронных цепей, образующих этот центр.
Свойства эти следующие:
1.Одностороннее проведение в нервных центрах можно доказать при раздражении передних корешков и отведении потенциалов от задних. В этом случае осциллограф не зарегистрирует импульсов. Если поменять электроды – импульсы будут поступать нормально.
2.Задержка проведения в синапсах. По рефлекторной дуге проведение возбуждения происходит медленнее, чем по нервному волокну. Это определяется тем, что в одном синапсе переход медиатора к постсинаптической мембране происходит за 0,3-0,5 мсек. (т.н. синаптическая задержка). Чем больше синапсов в рефлекторной дуге, тем больше время рефлекса, т.е. интервал от начала раздражения до начала деятельности. С учетом синаптической задержки проведение раздражения через один синапс требует около 1,5-2 мсек.
У человека наименьшую продолжительность имеет время сухожильных рефлексов (оно равно 20-24 мсек). У мигательного рефлекса оно больше – 50-200 мсек. Время рефлекса складывается из:
а) времени возбуждения рецепторов;
б) времени проведения возбуждения по центростремительным нервам;
в) времени передачи возбуждения в центре через синапсы;
г) времени проведения возбуждения по центробежным нервам;
д) времени передачи возбуждения на рабочий орган и латентного периода его деятельности.
Время “в” носит название центрального времени рефлекса.
Для упомянутых выше рефлексов оно составляет соответственно 3 мсек. и 36-180 мсек. Зная центральное время рефлекса, и учитывая, что через один синапс возбуждение проходит за 2 мсек., можно определить число синапсов в рефлекторной дуге. Например, коленный рефлекс считают моносинаптическим.
3. Суммация возбуждений. Впервые Сеченов показал, что в целостном организме рефлекторный акт может осуществляться при действии подпороговых стимулов, если они действуют на рецепторное поле достаточно часто. Такое явление получило название временной (последовательной) суммацией. Пример – рефлекс чесания у собаки можно вызвать, если подать в одну точку подпороговые стимулы с частотой 18 гц. Суммация подпороговых стимулов можно получить и тогда, когда они прикладываются на разные точки кожи, но одновременно – это пространственная суммация.
В основе этих явлений лежит процесс суммации возбуждающих постсинаптических потенциалов на теле и дендритах нейронов. При этом происходит накопление медиатора в синаптической щели. В естественных условиях оба вида суммации сосуществуют.
4. Центральное облегчение. Возникновение временной и особенно пространственной суммации способствуют и особенности организации синаптического аппарата в нервных центрах. Каждый аксон, поступая в ЦНС, ветвится и образует синапсы на большой группе нейронов (нейронный пул, или нейронная популяция). В такой группе принято условно различать центральную (пороговую) зону, и периферическую (подпороговую) кайму. Нейроны, находящиеся в центральной зоне, получают от каждого рецепторного нейрона достаточное количество синаптических окончаний для того, чтобы ответить разрядом ПД на приходящие импульсы. На нейронах же подпороговой каймы каждый аксон образует лишь небольшое число синапсов, возбуждение которых не способно возбудить нейрон. Нервные центры состоят из большого числа нейронных групп, причем отдельные нейроны могут входить в разные нейронные пулы. Это объясняется тем, что на одних и тех же нейронах оканчиваются разные афферентные волокна. При совместном раздражении этих афферентных волокон возбуждающие постсинаптические потенциалы в нейронах подпороговой каймы суммируются друг с другом и достигают критической величины. В результате в процесс возбуждения оказываются вовлеченными и клетки периферической каймы. При этом сила рефлекторной реакции суммарного раздражения нескольких “входов” в центр оказывается больше арифметической суммы раздельных раздражений. Этот эффект и носит название центрального облегчения.
5. Центральная окклюзия (закупорка). Может наблюдаться в деятельности нервного центра и обратный эффект, когда одновременное раздражение двух афферентных нейронов вызывает не суммацию возбуждения, а задержку, уменьшение силы раздражения. В этом случае суммарная реакция меньше арифметической суммы раздельных эффектов. Происходит это потому, что отдельные нейроны могут входить в центральные зоны разных нейронных популяций. В таком случае появление возбуждающих постсинаптических потенциалов на телах нейронов не приводят к увеличению числа
возбужденных одновременно клеток. Если суммация лучше проявляется при действии слабых афферентных раздражений, то явления окклюзии хорошо выражены с случае применения сильных афферентных раздражений, каждое их которых активирует большое число нейронов. Более наглядно эти эффекты видны на схемах.
6.Трансформация ритма возбуждений. Частота и ритм импульсов, поступающих к нервным центрам, и посылаемых ими на периферию, могут не совпадать. Это явление носит название трансформации. В ряде случаев на одиночный импульс, приложенный к афферентному волокну, мотонейрон отвечает серией импульсов. Образно говоря, в ответ на одиночный выстрел нервная клетка отвечает очередью. Чаще это бывает при длительном постсинаптическом потенциале и зависит от триггерных свойств аксонного холмика.
Центральное облегчение
Окклюзия
Рисунок 8. Схема возникновения центрального облегчения или окклюзии в нейронных популяциях.
Объяснение: Вверху- две нейронные популяции, у которых общими для разных аксонов является нейрон периферической каймы (3-й). При раздельном возбуждении 1 и 2-го аксонов возбуждается по 2 нейрона популяции (1,2 и 4.5 соответственно). При одновременно возбуждении этих аксонов возбуждается еще и 3 нейрон. Внизу- две нейронные популяции, у которых общими является нейрон центральной части (3). В этом случае и при раздельном возбуждении аксонов будут возбуждаться по 3 нейрона (1.2.3 и 3,4,5 соответственно), т.е. сумма отдельных возбуждений равна 6 нейронам. Однако при их одновременном возбуждении будут активироваться лишь те же 5 нейронов (1,2.3,4. и 5).
Другой механизм трансформации связан с эффектами сложения фаз двух или более волн возбуждения на нейроне – тут возможны эффекты как увеличения , так и снижения частоты выходящих из центра стимулов.
7. Последействие. Рефлекторные акты, в отличие от потенциалов действия, заканчиваются не одновременно с прекращением вызвавшего их раздражения, а через некоторый, иногда сравнительно длинный период времени. Продолжительность последействия может во много раз превышать продолжительность раздражения. Последействие обычно бывает больше при сильном и длительном раздражении.
Имеются два основных механизма, обусловливающих эффект последействия. Первый связан с суммацией следовой деполяризации мембраны при частых раздражениях (посттетаническая потенциация), когда нервная клетка продолжает давать разряды импульсов, несмотря на то, что кончилась серия раздражений. Второй механизм связывает последействие с циркуляцией нервных импульсов по замкнутым нейронным сетям рефлекторного центра.
8. Утомление нервных центров. В отличие от нервных волокон нервные центры легко утомляемы. Утомление нервного центра проявляется в постепенном снижении и в конечном итоге полном прекращении рефлекторного ответа при продолжительном раздражении афферентных нервных волокон. Если после этого приложить раздражение в эфферентному волокну – эффект возникает вновь.
Утомление в нервных центрах связано прежде всего с нарушением передачи возбуждения в межнейронных синапсах. Такое нарушение зависит от уменьшения запасов синтезированного медиатора, уменьшением чувствительности к медиатору постсинаптической мембраны, уменьшением энергетических ресурсов нервной клетки. Не все рефлекторные акты утомляются быстро (например, мало утомляемы проприоцептивные тонические рефлексы).
9. Рефлекторный тонус нервных центров. В его поддержании участвуют как афферентные импульсы, поступающие непрерывно от периферических рецепторов в ЦНС, так и различные гуморальные раздражители (гормоны, углекислота, и др.)
10. Высокая чувствительность к гипоксии. Показано, что 100 г. нервной ткани в единицу времени потребляет кислорода в 22 раза больше, чем 100 г. мышечной ткани. Поэтому нервные центры очень чувствительны к его недостатку. При этом чем выше центр, тем больше он страдает от гипоксии. Для коры мозга 5-6 минут достаточно, чтобы без кислорода произошли необратимые изменения, клетки ствола мозга выдерживают 15-20 минут полного прекращения кровообращения, а клетки спинного мозга – 20-30 минут. При гипотермии, когда снижается обмен веществ, ЦНС дольше переносит гипоксию.
11. Избирательная чувствительность к химическим веществам. Объясняется особенностями обменных процессов и позволяет находить фармпрепараты направленного действия.
Источник
Нервный центр – это совокупность нейронов, обеспечивающих регуляцию какого-либо конкретного физиологического процесса или функции.
Нервный центр в узком смысле – это совокупность нейронов, без которых данная конкретная функция не может регулироваться. Например, без нейронов дыхательного центра продолговатого мозга дыхание прекращается. Нервный центр в широком смысле — это совокупность нейронов, которые участвуют в регуляции конкретной физиологической функции, но не являются строго обязательными для ее осуществления! Например, в регуляции дыхания кроме нейронов продолговатого мозга участвуют нейроны пневмотаксического центра варолиевого моста, отдельные ядра гипоталамуса, кора больших полушарий и другие образования головного мозга.
Все нейроны нервного центра разделяют на 2 неравные по количеству и качеству группы.
Первая группа – нейроны центральной зоны. Это наиболее возбудимые нейроны, которые возбуждаются в ответ на поступление порогового (для нервного центра) сигнала. Таких нейронов около 15-20%, и они не обязательно располагаются в середине нервного центра, как это изображено на рис.1. Особенностью их является то, что они имеют на своем теле больше синаптических терминалей от сенсорных и вставочных нейронов.
Вторая группа – нейроны подпороговой каймы. Это менее возбудимые нейроны, которые не возбуждаются в ответ на поступление пороговых им-пульсов, но при действии более сильных раздражителей они возбуждаются и включаются в работу нервного центра, обеспечивая ее усиление. Таких нейронов большинство (80-85%), и они не обязательно располагаются на периферии нервного центра, но все имеют значительно меньше синаптических терминалей от сенсорных и вставочных нейронов по сравнению с нейронами центральной зоны.
На рис. 1 нейроны центральной зоны условно поставлены в центр внутреннего круга (А), а нейроны подпороговой каймы – в пространство между внутренним и наружным кругами (Б). Таким образом, если к нервному центру по афферентному входу (В) придет пороговый импульс, то возбудятся три нейрона центральной зоны, а на десяти нейронах подпороговой каймы потенциалы действия не возникнут, но появится местная деполяризация – возбуждающий постсинаптический потенциал (ВПСП).
От структуры нервного центра зависят его свойства, а они, в свою очередь, влияют на процесс проведения возбуждения через нервный центр, на его скорость и степень выраженности. От свойств нервных центров во многом зависит процесс распространения возбуждения по ЦНС, что имеет важное значение в интегративной деятельности организма.
Свойства нервных центров обусловлены описанной выше нейронной организацией нервного центра, а также химическим способом передачи возбуждения в синапсах. При электрическом способе передачи возбуждения нервные центры не имели бы подобных свойств.
Свойства нервных центров: 1 одностороннее проведение возбуждения; 2 задержка проведения возбуждения; 3 суммация; 4 облегчение; 5 окклюзия; 6 мультипликация; 7 трансформация; 8 последействие; 9 посттетаническая потенциация; 10 утомление; 11 тонус; 12 высокая чувствительность к изменению состояния внутренней среды организма; 13 пластичность.
1) Свойство «одностороннее проведение возбуждения» прямо связано со структурно-функциональными особенностями синапса. В синапсе медиатор выделяется из пресинаптического аппарата и поступает на постсинаптическую мембрану, на которой находятся белки-рецепторы, чувствительные к этому медиатору (они закрывают различные ионные каналы на постсинаптической мембране). Следовательно, возбуждение через синапс, а значит, и через нервный центр проходит только в одну сторону.
2) Свойство «задержка проведения возбуждения» также связано с химическим способом передачи возбуждения в синапсах. В отличие от электрического, при этом способе на передачу возбуждения в синапсе, а значит, и в нервном центре затрачивается больше времени (выделение медиатора из пресинаптического аппарата, поступление его на постсинаптическую мембрану, контакт с белками-рецепторами и т.д.), чем на проведение возбуждения по нервному волокну. Русский физиолог А.Ф. Самойлов (1924) определил, что скорость проведения возбуждения по нервному волокну в 1,5 раза больше, чем через синапс. На основании этого факта ученый высказал предположение, что в основе проведения возбуждения по нервному волокну лежат физические процессы, а в основе синаптического способа передачи – химические.
Время проведения возбуждения («синаптическая задержка») через синапсы соматической нервной системы составляет 0,5-1 мс, а через синапсы вегетативной нервной системы – до 10 мс.
3) Суммация – это возникновение возбуждения в нервном центре при поступлении к нему нескольких допороговых импульсов, каждый из которых в отдельности не может возбуждения (рис. 2). Фактически этот процесс происходит на нейронах подпороговой каймы. Различают два вида суммации: пространственную и временною.
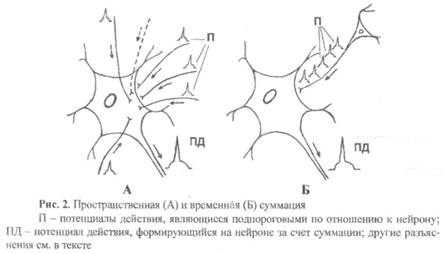 Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Пространственная суммация возникает в том случае, когда к нервному центру (к его нейронам) приходят одновременно, несколько допороговых импульсов. На рисунке 2А видно, что к нейрону подпороговой каймы, имеющему пороговый потенциал 30 мВ одновременно по пяти различным афферентным входам (их аксоны обозначены сплошной линией) приходят пять импульсов, каждый из которых деполяризует мембрану нейрона на 5 мВ (то есть возникают пять отдельных ВПСП). В этом случае возбуждение нейрона не наступает, так как суммарная деполяризация мембраны нейрона составляет лишь 25 мВ (суммированный ВПСП мал для достижения КУД). Но если к нейрону придет еще один подобный импульс по шестому входу (его аксон обозначен пунктирной линией), то суммированный ВПСП будет достаточен по величине и мембрана нейрона в зоне аксонного холмика деполяризуется до критического уровня, в результате чего нейрон из состояния покоя перейдет в состояние возбуждения. На постсинаптической мембране происходит суммация ВПСП в пространстве.
Временная (последовательная) суммация возникает в том случае, когда к нейронам нервного центра по одному афферентному входу приходит не один, а серия импульсов с очень небольшими по времени межимпульсными промежутками (рис. 2Б). Два механизма временной суммации:
1) интервалы между отдельными импульсами настолько малы, что за это время медиатор, выделившийся в синаптическую щель, не успевает полностью разрушиться и вернуться в пресинаптический аппарат. В этом случае возникает постепенное накопление медиатора до критического объема, необходимого для возникновения достаточного по амплитуде ВПСП, а значит, и для возникновения ПД;
 2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
2) интервалы между отдельными импульсами настолько малы, что возникший за это время на постсинаптической мембране ВПСП не успевает исчезнуть и усиливается за счет новой порции медиатора – суммируется. На постсинаптической мембране происходит суммация ВПСП во времени.
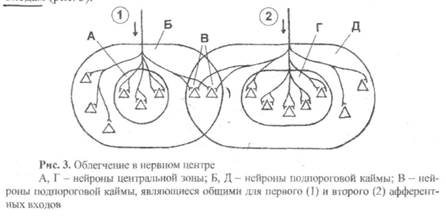
4) Облегчение – это увеличение количества возбужденных нейронов в нервном центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному, а по двум или более афферентным входам. На рис. 3 рассмотрен случай, когда при отдельном раздражении первого афферентного входа возбуждается только три нейрона центральной зоны (А), а на пяти нейронах подпороговой каймы (Б) возникают ВПСП. Если раздражать отдельно только второй афферентный вход, то возбуждены будут пять нейронов (Г), а четыре нейрона подпороговой каймы (Д) не возбудятся. Раздражая и первый, и второй афферентные входы одновременно (!), мы ожидаем вовлечения в процесс возбуждения восьми нейронов. И они, естественно, будут возбуждаться, но кроме них (сверх ожидания!) могут возбуждаться еще некоторые нейроны подпороговой каймы. Это произойдет потому, что один или несколько нейронов подпороговой каймы являются общими как для первого, так и для второго афферентных входов (в нашем случае это два нейрона, обозначенные буквой В), и при одновременном поступлении возбуждения к этим нейронам дни возбудятся за счет возникновения пространственной суммации.
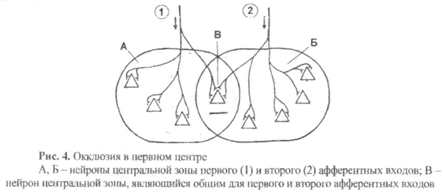 5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
5) Окклюзия – это уменьшение количества возбужденных нейронов в нервной центре (по сравнению с ожидаемым) при одновременном поступлении к нему возбуждения не по одному. а по двум или более афферентным входам (рис. 4).
 На рис. 4 видно, что при поступлении возбуждения только по первому афферентному входу возбуждаются четыре нейрона, а при раздражении только второго афферентного входа – пять нейронов, так как и в том, и другом случае они относятся к центральным зонам. Понятно, что при одновременном поступлении возбуждения по первому и второму входам мы ожидаем увидеть девять возбужденных нейронов, но на самом деле таких нейронов будет только восемь. Это произойдет потому, что нейрон, обозначенный буквой В, является общим для обоих входов и по закону «все или ничего» будет возбуждаться в любом случае независимо от того, сколько пороговых импульсов к нему прилет одновременно.
На рис. 4 видно, что при поступлении возбуждения только по первому афферентному входу возбуждаются четыре нейрона, а при раздражении только второго афферентного входа – пять нейронов, так как и в том, и другом случае они относятся к центральным зонам. Понятно, что при одновременном поступлении возбуждения по первому и второму входам мы ожидаем увидеть девять возбужденных нейронов, но на самом деле таких нейронов будет только восемь. Это произойдет потому, что нейрон, обозначенный буквой В, является общим для обоих входов и по закону «все или ничего» будет возбуждаться в любом случае независимо от того, сколько пороговых импульсов к нему прилет одновременно.
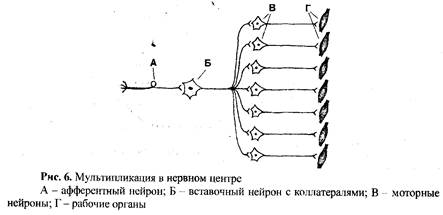
6) Мультипликационное возбуждение (мультипликация) заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение поступает одновременно не на один, а на несколько моторных нейронов (рис. 6). В связи с этим эффект на рабочем органе усиливается в несколько раз, или в работу вовлекаются не одна, а несколько рабочих структур, Это свойство особенно ярко проявляется в ганглиях автономной (вегетативной) нервной системы.
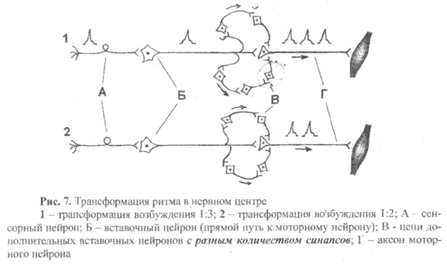 7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
7) Трансформация ритма возбуждения – это изменение частоты импульсов на выходе из нервного центра по сравнению с частотой импульсов на входе в нервный центр.
Частота импульсов на выходе из нервного центра может быть значительно меньше, чем на входе. Говоря техническим языком, это «понижающая трансформация». Подобное явление мы уже рассматривали выше («временная суммация»).
Частота импульсов на выходе из нервного центра может быть значительно выше, чем на входе («повышающая трансформация»). Это связано с особенностями взаимосвязи вставочных нейронов:
а) наличием дублирующих цепей вставочных нейронов, связывающих сенсорные и моторные нейроны;
б) разным количеством синапсов в каждой из этих цепей.

 Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Например, на рис.7 представлены два варианта трансформации, которые, на первый взгляд, не отличаются друг от друга, так как в том и в другом случае показаны две дополнительные цепи вставочных нейронов (кроме прямого пути), с помощью которых возбуждение может передаваться по цепи нейронов А—Б—В. Рассмотрим эти схемы.
Вариант 1. Верхняя цепь состоит из двух дополнительных вставочных нейронов, а значит, по сравнению с прямым путем передачи возбуждения с нейрона Б на нейрон В, имеет два дополнительных синапса. Поэтому возбуждение, проходя по верхней цепи, задержится на 2 мс (время синаптической задержки в одном синапсе составляет ~1 мс) и придет на нейрон В после того, как пройдет возбуждение по прямому пути. В нижней цепи три дополнительных вставочных нейрона (то есть три дополнительных синапса), значит, возбуждение будет доходить до нейрона В еще дольше, чем по верхней цепи (задержка составит 3 мс). Следовательно, по нижней цепи возбуждение на нейрон В придет после того, как пройдет возбуждение по верхней цепи. В результате на один импульс, пришедший по сенсорному нейрону А, на моторном нейроне В возникнет три потенциала действия (трансформация 1:3).
Вариант 2. В этом случае и верхняя и нижняя цепи вставочных нейронов состоят из двух дополнительных нейронов. Возбуждение по обеим цепям придет к нейрону В одновременно в виде одного потенциала действие, который появится на нейроне В только после прохождения возбуждения к нему от нейрона Б по прямому пути. В этом варианте мы тоже получим трансформацию ритма, но уже в соотношении 1:2.
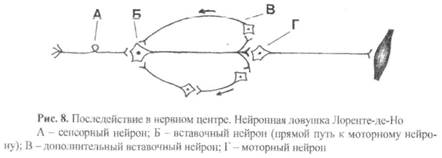 8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
8) Последействие – это продолжение возбуждения моторного нейрона в течение некоторого времени после прекращения действия раздражителя.
Сущность механизма последействия заключается в том, что по разветвлениям аксона вставочного нейрона возбуждение распространяется на соседние вставочные нейроны и по ним возвращается на первоначальный вставочный нейрон. Возбуждение как бы «запирается» в нейронной ловушке и циркулирует в ней достаточно долго (рис. 8). Наличием таких нейронных ловушек объясняют, в частности, механизм кратковременной памяти.
Другими причинами последействия могут быть:
а) возникновение высокоамплитудного ВПСП, в результате которого возникает не один, а несколько потенциалов действия то есть ответ длится большее время;
б) длительная следовая деполяризация постсинаптической мембраны, в результате чего возникают несколько потенциалов действия, вместо одного.
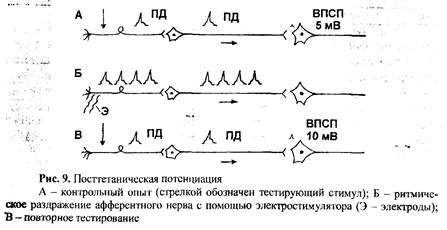 9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
9) Посттетаническая потенциация (синаптическое облегчение) – это улучшение проведения в синапсах после короткого раздражения афферентных путей.
Если в качестве контроля вызвать одиночное раздражение афферентного нерва тестирующим раздражителем (рис. 9А), то на моторном нейроне мы получим ВПСП вполне определенной амплитуды (в нашем случае 5 мВ). Если после этого тот же афферентный нерв раздражать некоторое время серией частых импульсов (рис. 9Б), а потом вновь подействовать тестирующим раздражителем (рис. 9В), то величина ВПСП будет больше (в нашем случае 10 мВ). Причем она будет тем больше, чем более частыми импульсами мы раздражали афферентный нерв.
Длительность синаптического облегчения зависит от свойств синапса и характера раздражения: после одиночных стимулов оно выражено слабо, после раздражающей серии потенциация (облегчение) может продолжаться от нескольких минут до нескольких часов. Объясняется он тем, что при частом раздражении афферентного волокна в его пресинаптической терминали (окончании) накапливаются ионы кальция, а значит, улучшается выделение медиатора. Кроме того, показано, что частое раздражение нерва приводит к усилению синтеза медиатора, мобилизации пузырьков медиатора, к усилению синтеза белков-рецепторов на постсинаптической мембране и увеличению их чувтствительности. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах.
10) Утомление нервного центра (посттетаническая депрессия, синаптическая депрессия) – это уменьшение или прекращение импульсной активности нервного центра в результате длительной стимуляции его афферентными импульсами (или произвольного вовлечения его в процесс возбуждения посредством импульсов, идущих из коры больших полушарий). Причинами утомления нервного центра могут быть:
• истощение запасов медиатора в афферентном или вставочном нейроне;
• снижение возбудимости постсинаптической мембраны (то есть мембраны моторного или вставочного нейрона) из-за накопления, например, продуктов метаболизма.
Утомляемость нервных центров продемонстрировал Н.Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения п. tibialis и п. peroneus. В этом случае ритмическое раздражение одного нерва вызывает ритмические сокращения мышцы, приводящие к ослаблению силы ее сокращения вплоть до полного отсутствия сокращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локализации утомления не в мышце, а в центральной части рефлекторной дуги. Синаптическая депрессия при длительной активации центра выражается в снижении постсинаптических потенциалов.
11) Тонус нервного центра – это длительное, умеренное возбуждение нервного центра без видимо утомления Причинами тонуса могут быть:
• потоки афферентных импульсов, постоянно поступающие с неадаптирующихся рецепторов;
• гуморальные факторы, постоянно присутствующие в плазме крови;
• спонтанная биоэлектрическая активность нейронов (автоматия);
• циркуляция (реверберация) импульсов в ЦНС.
12) Нервный центр состоит из нейронов, а они очень чувствительны к изменению состава внутренней среды организма, что и отражается на свойствах нервных центров. Наиболее важными факторами, влияющими на работу нервных центров, являются: гипоксия; недостаток питательных веществ (например, глюкозы); изменение температуры; воздействие продуктов метаболизма; воздействие различных токсических и фармакологических препаратов.
Разные нервные центры имеют неодинаковую чувствительность к воздействию названных факторов. Так, нейроны коры больших полушарий наиболее чувствительны к гипоксии, недостатку глюкозы, продуктам метаболизма; клетки гипоталамуса – к изменению температуры, содержанию глюкозы, аминокислот, жирных кислот и др.; различные участки ретикулярной формации выключаются разными фармакологическими препаратами, различные нервные центры избирательно активируются или тормозятся разными медиаторами.
13) Пластичность нервного центра означает его способность изменять при определенных обстоятельствах свои функциональные свойства. В основе этого явления лежит поливалентность нейронов нервных центров. Особенно ярко проявляется это свойство при всевозможных повреждениях ЦНС, когда организм компенсирует утраченные функции за счет сохранившихся нервных центров. Особенно хорошо свойство пластичности выражено в коре больших полушарий. Например, центральные параличи, связанные с патологией двигательных центров коры, иногда полностью компенсируются, и ранее утраченные двигательные функции восстанавливаются.
Источник