
Какими свойствами обладает постсинаптическая мембрана

Субсинаптическая мембрана – участок постсинаптической мембраны, на котором её свойства выражены максимально.
Свойства субсинаптической мембраны :
А) она хемовозбудима.
Б) она электроневозбудима
в) она имеет большое число однотипных хеморецепторов, которые воспринимают действие медиатора и высокую концетрацию соответствующих ионных каналов (хемочувствительные,рецепторуправляемые каналы)
Синаптическая щель.
Размер 200-500 ангстрем /20-50 мкм (микрон)/, заполнена межклеточной жидкостью, существует периферический барьер, /что это такое никто не знает/ препятствующий выходу медиатора за пределы синапса.
Механизм передачи возбуждения через синапс
Когда возбуждение приходит в область пресинаптической мембраны, она деполяризуется, это:
Активирует кальциевые потенциалзависимые каналы N-типа, они медленные, быстро инактивируются. В клетку входит небольшое количество кальция
Вошедший кальций связывается со специальным белком (кальций – связывающий белок) – синаптосин. Кальций активирует фосфолирирование нескольких молекул синаптосина, вызывая их конформацию, что приводит к ослаблению связей везикул с цитоскелетом клетки и они (везикулы) перемещаются к внутренней поверхности пресинаптической и прилипает к ней. Это вызывает разрыв везикул (ферментативный протеолиз). Параллельно прилипание к мембране активирует мембранный белок- синаптопор, который выступает как переносчик медиатора через мембрану либо как белок – переносчик за счет конформации молекулы, либо образуя транспортный канал, либо вызывая экзоцитоз.
Медиатор диффундирует в синаптическую щель, он не выходит за пределы синаптической щели, т.к. сбоку имеются синаптические барьеры, которые препятствуют выходу медиатора за пределы синапса (что такое синаптические барьеры – неизвестно). Молекулы медиатора выделяются квантами. Количество выделившихся квантов зависит от силы и частоты раздражения, т.е. передача возбуждения в синапсе за счёт этого квантируется, она становится дискретной.
Молекулы медиатора идут в к постсинаптической мембране, в область субсинаптической мембраны, которая имеет много однотипных хеморецепторов и образуют комплекс «медиатор – рецептор». Это вызывает активацию соответствующих рецепторуправляемых ионных каналов.
Все медиаторы можно разделить на возбуждающие медиаторы и тормозные медиаторы. Следовательно и синапсы делятся на возбуждающие и тормозные.
Возбуждающие медиаторы взаимодействуя с рецептором субсинаптической мембраны вызывают активацию натриевых каналов и формируют входящий натриевый ток, который вызывает возникновение частичной деполяризации, т.е рецепторный потенциал, который на уровне синапс обозначают как возбуждающий постсинаптический потенциал (ВПСП). Тормозной медиатор вызывает усиление входящего калиевого тока или входящего ток хлора, т.е. вызывает локальную гиперполяризацию. Это формирует тормозной постсинаптический потенциал (ТПСП). Рисунок. Конечный эффект (потенциал действия или тормозной потенциал) формируется за счет суммации ВПСП или ТПСП.
В синапсе известно два вида суммации:
1. Пространственная суммация локальных очагов возбуждения (когда в пространстве суммируются эти локальные очаги и возникает потенциал действия);
2. Временная суммация (когда на одни и те же рецепторы последовательно через короткие интервалы времени воздействуют новые порции медиатора и возникает как бы ступенечка – частичная деполяризация, она не окончилась, на нее наслаивается следующая частичная деполяризация и так поляризация достигает КУДа – временная суммация)
В дальнейшем комплекс «медиатор-рецептор» диссоциирует. Если этого не происходит, то в возбуждающих синапсах возникает стойкая деполяризация по типу катодической депрессии, При этом данный рецептор перестает воспринимать какую-либо другую информацию.
Поэтому в нормальных, естественных условиях медиатор отсоединяется от рецепторов и разрушается ферментами (холинэстераза и т.д.), которые имеются в синапсе. Примерно 20-30% медиатора удаляется таким образом из синаптической щели.
Источник
Медиатор пресинаптической мембраны. Постсинаптическая мембранаМембрану пресинаптической терминали называют пресинаптической мембраной. Она содержит большое количество электроуправляемых кальциевых каналов. Когда потенциал действия деполяризует пресинаптическую мембрану, эти кальциевые каналы открываются, что позволяет большому числу ионов кальция войти в терминалы Количество медиатора, которое затем выделится из терминали в синаптическую щель, находится в прямой зависимости от числа вошедших ионов кальция. Считают, что ионы кальция, войдя в пресинаптическую терминаль, связываются с особыми белковыми молекулами на внутренней поверхности пресинаптической мембраны, которые называют местами освобождения. Это связывание вызывает открытие мест освобождения медиатора сквозь мембрану, в результате после каждого одиночного потенциала действия несколько пузырьков выделяют свой медиатор в щель. В каждом из пузырьков, содержащих медиатор ацетилхолин, накапливается от 2000 до 10000 его молекул, а общего числа пузырьков в пресинаптической терминали достаточно, чтобы проводить от нескольких сотен до более 10000 потенциалов действия. Действие медиатора на постсинаптический нейрон – функция рецепторных белков Мембрана постсинаптического нейрона содержит большое количество рецепторных белков. Молекулы этих рецепторов имеют два важных компонента:
Эти вещества, в свою очередь, служат вторичными посредниками для усиления или угнетения специфических клеточных функций. Катионные каналы, проводящие ионы Na+, выстланы отрицательными зарядами. Эти заряды затягивают положительно заряженные ионы Na+ в канал, если диаметр канала превышает размер гидратированного иона Na+. Однако те же самые отрицательные заряды отталкивают ионы Сl- и другие анионы, препятствуя их прохождению. Если диаметры анионных каналов достаточно велики, ионы Сl- входят в них и проходят на противоположную сторону, тогда как вход натриевых, калиевых и кальциевых катионов блокируется в основном из-за слишком больших размеров их гидратированных ионов. Далее мы узнаем, что процесс открытия катионных каналов, позволяющий войти в клетку положительно заряженным ионам Na+, способствует возбуждению нейрона, поэтому медиаторы, ведущие к открытию катионных каналов, называют возбуждающими. И наоборот, процесс открытия анионных каналов, позволяющий войти отрицательным электрическим зарядам, тормозит нейрон. Соответственно, медиаторы, открывающие эти каналы, называют тормозными. При активации медиатором ионного канала он обычно открывается в течение долей миллисекунды; после удаления медиатора канал закрывается так же быстро. Такое открытие и закрытие каналов обеспечивает очень быстрое управление постсинаптическими нейронами. – Также рекомендуем “Вторичные посредники синапса. Рецепторы постсинаптической мембраны” Оглавление темы “Нервная система и его физиология”: |

Источник
Постсинаптической мембраны
Синаптической щели
Пресинаптическая мембрана является окончанием отростка нервной клетки. Внутри отростка в непосредственной близости от мембраны имеется скопление пузырьков (гранул), содержащих тот или иной медиатор. Пузырьки находятся в постоянном движении.
Постсинаптическая мембрана является частью клеточной мембраны иннервируемой ткани. Постсинаптическая мембрана в отличие от пресинаптической имеет белковые хеморецепторы к биологически активным (медиаторам, гормонам), лекарственным и токсическим веществам. Важная особенность рецепторов постсинаптической мембраны – их химическая специфичность, т.е. способность вступать в биохимическое взаимодействие только с определенным видом медиатора.
Синаптическая щель представляет собой пространство между пре- и постсинаптичекой мембранами, заполненное жидкостью, близкой по составу к плазме крови. Через нее медиатор медленно диффундирует от пресинаптической мембраны к постсинаптической.
Особенности строения нервно-мышечного синапса обусловливают его физиологические свойства.
Одностороннее проведение возбуждения (от пре- к постсинаптической мембране), обусловленное наличием чувствительных к медиатору рецепторов только в постсинаптической мембране.
Синаптическая задержка проведения возбуждения (время между приходом импульса в пресинаптическое окончание и началом постсинаптического ответа), связанная с малой скоростью диффузии медиатора в синаптическую щель по сравнению со скоростью прохождения импульса по нервному волокну.
Низкая лабильность и высокая утомляемость синапса, обусловленная временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности.
Высокая избирательная чувствительность синапса к химическим веществам, обусловленная специфичностью хеморецепторов постсинаптической мембраны.
Этапы синаптической передачи.
Синтез медиатора. В цитоплазме нейронов и нервных окончаний синтезируются химические медиаторы – биологически активные вещества. Они синтезируются постоянно и депонируются в синаптических пузырьках нервных окончаний.
Секреция медиатора. Высвобождение медиатора из синаптических пузырьков имеет квантовый характер. В состоянии покоя оно незначительно, а под влиянием нервного импульса резко усиливается.
Взаимодействие медиатора с рецепторами постсинаптической мембраны. Это взаимодействие заключается в избирательном изменении проницаемости ионоселективных каналов эффекторной клетки в области активных центров связывания с медиатором. Взаимодействие медиатора со своими рецепторами может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. В случае увеличения проницаемости натриевых и кальциевых каналов усиливается поступление Na и Ca в клетку с последующей деполяризацией мембраны, возникновением ПД и дальнейшей передачей нервного импульса. Такие синапсы называются возбуждающими. Если повышается проницаемость калиевых каналов и каналов для хлора, наблюдается избыточный выход К из клетки с одновременной диффузией в нее Cl , что приводит к гиперполяризации мембраны, снижению ее возбудимости и развитию тормозных постсинаптических потенциалов. Передача нервных импульсов затрудняется или совсем прекращается. Такие синапсы называются тормозными.
Рецепторы, взаимодействующие с АХ, называются холинорецепторами. В функциональном отношении они разделяются на две группы: М – и Н-холинорецепторы. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, тогда как в мышцах внутренних органов – преимущественно М-холинорецепторы.
Рецепторы, взаимодействующие с НА, называются адренорецепторами. В функциональном отношении они делятся на альфа- и бета-адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие НА является деполяризующим, если он взаимодействует с альфа-адренорецепторами (сокращение мышечной оболочки стенок кровеносных сосудов или кишечника), или тормозным – при взаимодействии с бета-адренорецепторами (их расслабление).
Инактивация медиатора. Инактивация (полная потеря активности) медиатора необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня МП. Наиболее важным путем инактивации является гидролитическое расщепление с помощью ингибиторов. Для АХ ингибитором является холинэстераза, для НА и адреналина – моноаминоксидаза и катехолоксиметилтрансфераза.
Другой путь удаления медиатора из синаптической щели – «обратный захват» его пресинаптическими окончаниями (пиноцитоз) и обратный аксонный транспорт, особенно выраженный для катехоламинов.
В основе координационной деятельности ЦНС лежит взаимодействие процессов возбуждения и торможения.
Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани.
Торможение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся снижением функций ткани.
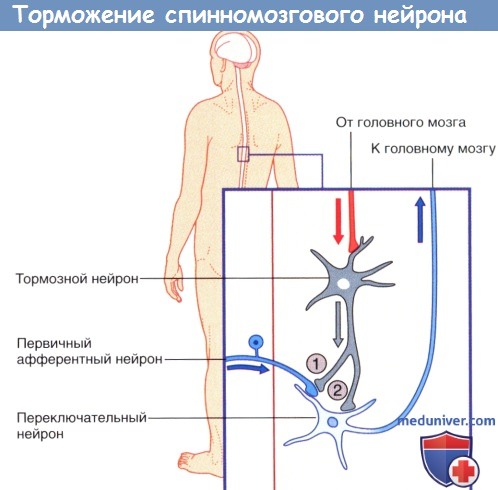
Первичное торможение в ЦНС возникает за счет тормозных нейронов. Это особый вид вставочных нейронов, которые при передаче импульса выделяют тормозной медиатор. Различают 2 вида первичного торможения: постсинаптическое и пресинаптическое.
Постсинаптическое торможение возникает, если аксон тормозного нейрона образует синапс с телом нейрона и, выделяя медиатор, вызывает гиперполяризацию клеточной мембраны, тормозя активность клетки.
Пресинаптическое торможение возникает, когда аксон тормозного нейрона образует синапс с аксоном возбуждающего нейрона, препятствуя проведению импульса.
Читайте также:
Рекомендуемые страницы:
©2015-2020 poisk-ru.ru
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Дата создания страницы: 2016-02-12
Нарушение авторских прав и Нарушение персональных данных
Источник
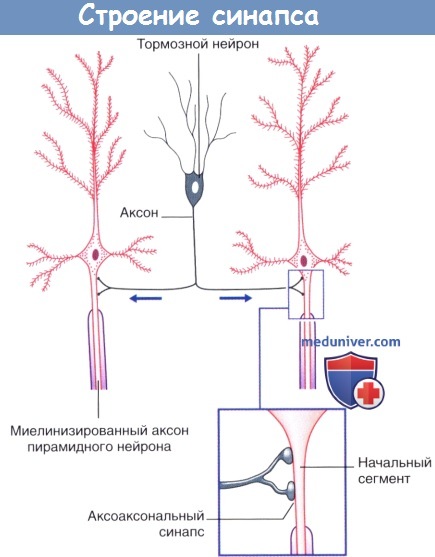
Строение синапса: электрические и химические синапсыОбласть контакта между двумя нейронами называют синапсом.
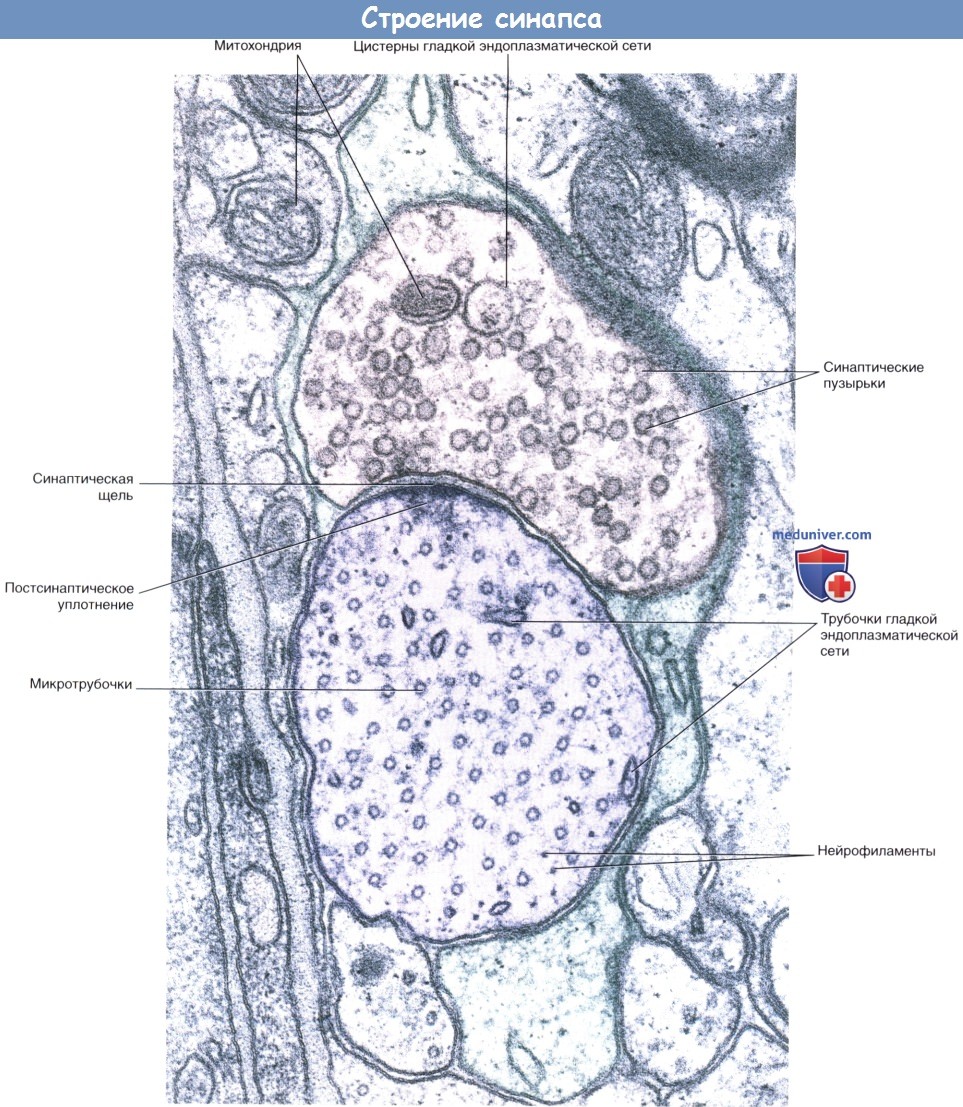
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов. Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому. б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал. Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы. В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
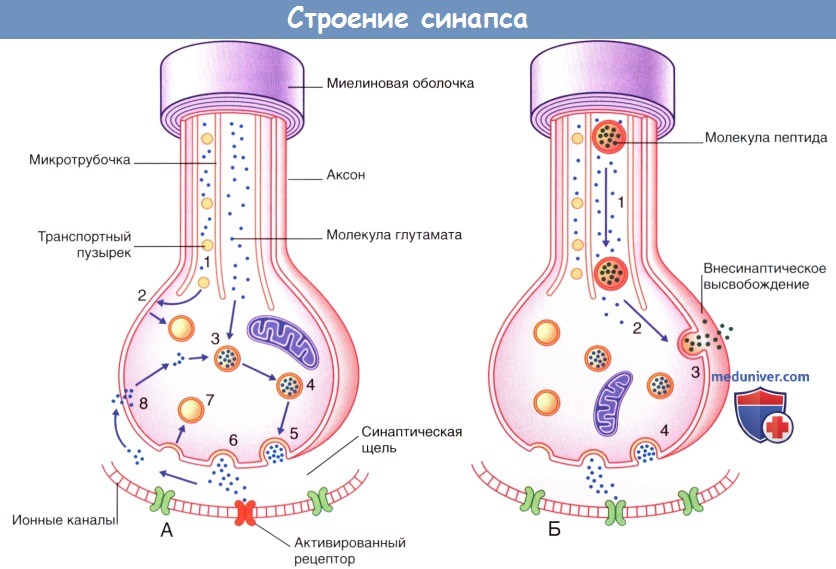
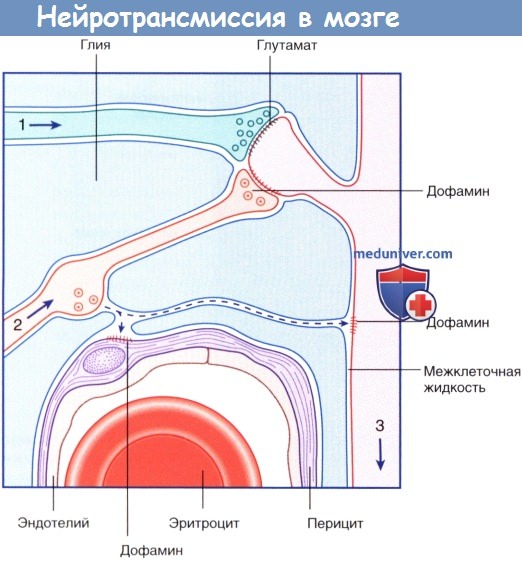
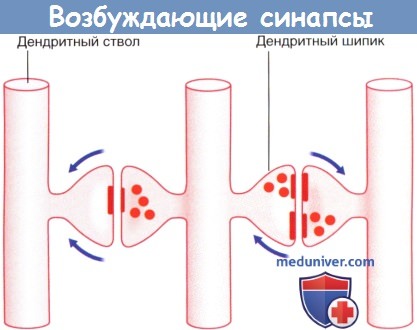
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона. Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия. Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат. Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем. Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата. Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора. Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию. Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором. В отличие от диффузной нейротрансмиссии, традиционную синаптическую передачу сигнала ввиду ее относительной стабильности называют «проводниковой». в) Резюме. Мультиполярные нейроны ЦНС состоят из сомы, дендритов и аксона; аксон образует коллатеральные и терминальные ветви. В соме расположены гладкая и шероховатая эндоплазматическая сети, комплексы Гольджи, нейрофиламенты и микротрубочки. Микротрубочки пронизывают нейрон на всем протяжении, принимают участие в процессе антероградного транспорта синаптических пузырьков, митохондрий и веществ для построения мембран, а также обеспечивают ретроградный транспорт «маркерных» молекул и разрушенных органелл. Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое). Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной. Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
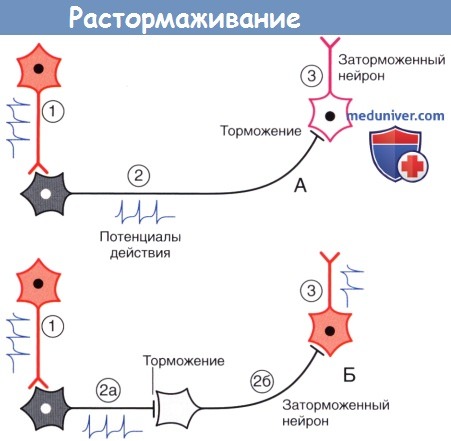
2. Лекарственные средства – «ключи» и «замки». Рецептор можно сравнить с замком, а медиатор — с подходящим к нему ключом. В том случае, если процесс высвобождения медиатора нарушится с возрастом или в результате какого-либо заболевания, лекарственное средство может сыграть роль «запасного ключа», выполняющего аналогичную медиатору функцию. Такое лекарственное средство называют агонистом. В то же время в случае чрезмерной продукции медиатор может быть «перехвачен» блокатором рецептора — «фальшивым ключом», который свяжется с «замком»-рецептором, но при этом не вызовет его активацию. 3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях. 4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение. Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса. Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
Учебное видео – строение синапса– Также рекомендуем “Нейроглиальные клетки ЦНС. Клетки нейроглии” Редактор: Искандер Милевски. Дата публикации: 11.11.2018 |
Источник