
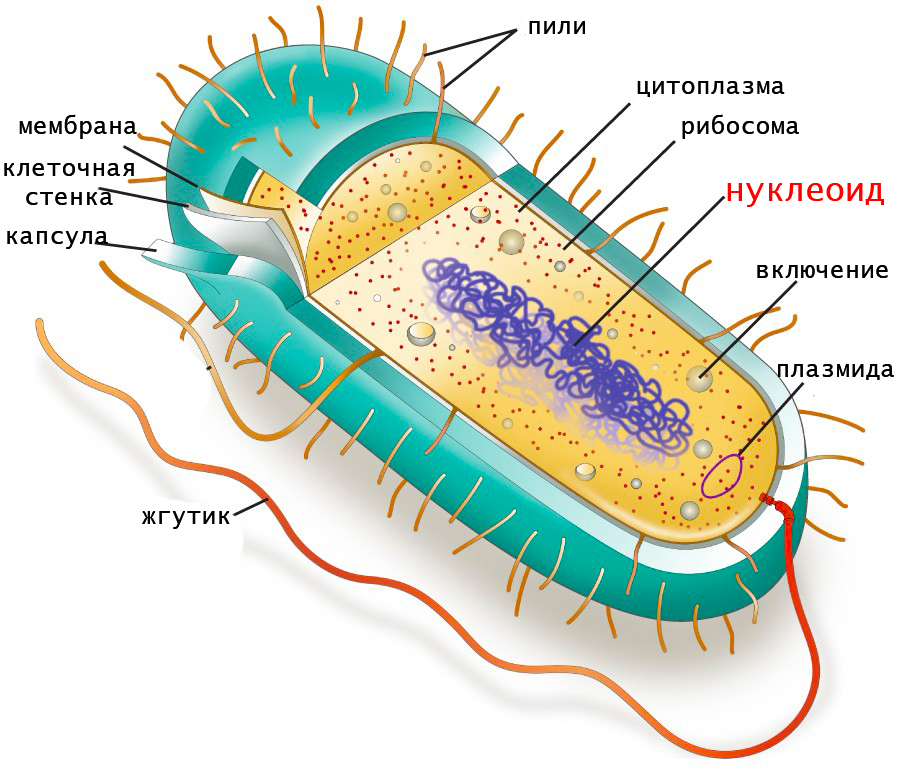
В каких клетках содержится нуклеоид

Микрофотография бактериальной клетки с нуклеоидом, выделенным зелёной пунктирной линией
Нуклео́ид (англ. Nucleoid) — неправильной формы зона в цитоплазме прокариотической клетки, в которой находится геномная ДНК и ассоциированные с ней белки. На долю ДНК приходится около 60 % массы нуклеоида; помимо ДНК, нуклеоид содержит РНК и белки[1]. Белки нуклеоида, которые обеспечивают пространственную организацию геномной ДНК, называют нуклеоидными белками или нуклеоид-ассоциированными белками; они не имеют ничего общего с гистонами, упаковывающими ДНК у эукариот. В отличие от гистонов, ДНК-связывающие белки[en] нуклеоида не формируют нуклеосомы и обеспечивают компактизацию геномной ДНК другим способом[2]. Несмотря на аморфную форму, отдельные гены располагаются в нём упорядоченно[3].
Бактерии[править | править код]
Бактериальные хромосомы[править | править код]
В большинстве случаев геном бактерии представлен кольцевой молекулой ДНК[en], которую также называют хромосомой. Как правило, бактериальная хромосома имеет длину около 1 мм, она занимает до 20 % цитоплазмы и имеет объём около 0,2 мкм³. Репликация геномной ДНК начинается от участка инициации (oriC), из которого две репликативные вилки движутся в противоположных направлениях и встречаются в сайте терминации (Ter), в котором далее происходит размыкание сцепленных дочерних хромосом. Количество хромосом в бактериальной клетке зависит не только от видовой принадлежности, но и от фазы развития популяции. В качестве бактерий, постоянно имеющих несколько хромосом, можно привести Deinococcus radiodurans (от 4 до 10 хромосом), Borrelia hermsii[en] (от 8 до 16 хромосом), Desulfovibrio gigas (от 9 до 17 хромосом), Azotobacter vinelandii (до 80 хромосом). Молодые клетки обычно содержат больше хромосом, чем старые. Иногда хромосомы представляют не просто копии геномной ДНК: в некоторых случаях геном распределяется между несколькими неодинаковыми хромосомами и внехромосомными элементами (плазмидами). Так, у Agrobacterium tumefaciens, Brucella melitensis[en] и Rhodobacter sphaeroides[en] имеются две разные хромосомы, а у Bacillus cereus, Leptospira interrogans[en] и Rhizobium meliloti[en] кроме хромосом есть одна или две мегаплазмиды, состоящие из 100—500 тысяч пар оснований (п. о.)[4].
Ряд бактерий имеют и кольцевые, и линейные хромосомы, а некоторые — только линейные. Например, линейная хромосома имеется у возбудителя болезни Лайма — Borrelia burgdorferi[en]. Известны и линейные плазмиды. Роль теломер при этом могут выполнять или одноцепочечные концевые шпильки, или особые белки, ковалентно связанные с концами линейной хромосомы или плазмиды; в некоторых случаях на время репликации линейная хромосома становится кольцевой[5].
ДНК-связывающие белки[править | править код]
За компактизацию бактериальной хромосомы отвечают несколько разнородных белков, из которых наиболее важны HU, H-NS и SMC. Белки HU и H-NS называют гистоноподобными, их взаимодействие с ДНК зависит от её последовательности, наличия в молекуле шпилек и изгибов. Выявлены также минорные гистоноподобные белки FIS и IHF, которые, наряду с HU и H-NS, не только обеспечивают пространственную организацию бактериальной геномной ДНК, но также влияют на её репликацию, рекомбинацию и экспрессию генов. HU, совместно с ДНК-гиразой, обеспечивает отрицательную сверхспирализацию ДНК. H-NS компактизует ДНК, влияет на её сверхспирализацию, вызывает в ней изгибы, однако по большей части он задействован в экспрессии генов: под его контролем находятся до 40 генов. Белки SMC имеются у представителей всех трёх доменов жизни. Они гомологичны миозину, у эукариот они играют роль моторов при конденсации хроматина[6].
Таким образом, можно выделить следующие уровни компактизации бактериальной ДНК. Первый уровень обеспечивается гистоноподобными белками HU и H-NS. Второй уровень — отрицательная сверхспирализация — обеспечивается SMC-белками и топоизомеразами. Наконец, высший уровень компактизации представлен независимо сверхспирализованными петлями (доменами), на которые разделена бактериальная хромосома. В состав каждого домена входит около 10 тысяч п. о[7].
Археи[править | править код]
У архей компактные нуклеоиды располагаются в центре клетки, как и у бактерий, и ориентированы вдоль её продольной оси. Число хромосом зависит от видовой принадлежности и фазы развития популяции, кроме того, в большинстве случаев в молодых клетках содержится больше хромосом. Так, у Methanocaldococcus jannaschii[en] в стационарной фазе роста имеется от одной до пяти хромосом, а в фазе экспоненциального роста — 3—15 хромосом. Как и бактерии, археи могут иметь плазмиды[8].
У многих архей, например, у эвриархеот, нижний уровень компактизации ДНК обеспечивается белками, гомологичными гистонам эукариот. Гистоны архей по размеру уступают гистонам эукариот и имеют укороченный N-концевой участок, поэтому взаимодействуют с ДНК несколько иначе. Нуклеосомы архей не имеют октамерной структуры в силу отсутствия гистонов H2A[en] и H2B[en], однако они похожи на тетрасому (H3[en]/H4[en])2. Кренархеоты, ведущие термофильный и гипертермофильный образ жизни, не имеют гистонов, но у них имеются негистоновые ДНК-связывающие белки. Например, один из них, Sul7D, взаимодействует с малой бороздкой ДНК и защищает её от тепловой денатурации[9].
В отличие от бактерий, геномная ДНК архей характеризуется положительной сверхспирализацией, которая, как полагают, стабилизирует ДНК у экстремофильных видов. Положительную сверхспирализацию обеспечивает так называемая «обратная» гираза[10].
Примечания[править | править код]
- ↑ Нетрусов, Котова, 2012, с. 37—38.
- ↑ Wang W., Li G. W., Chen C., Xie X. S., Zhuang X. Chromosome organization by a nucleoid-associated protein in live bacteria. (англ.) // Science (New York, N.Y.). — 2011. — 9 September (vol. 333, no. 6048). — P. 1445—1449. — doi:10.1126/science.1204697. — PMID 21903814. [исправить]
- ↑ Кассимерис, Лингаппа, Плоппер, 2016, с. 945.
- ↑ Пиневич, 2006, с. 194—195.
- ↑ Пиневич, 2006, с. 195.
- ↑ Пиневич, 2006, с. 196—197.
- ↑ Пиневич, 2006, с. 197—198.
- ↑ Пиневич, 2006, с. 198.
- ↑ Пиневич, 2006, с. 198—199.
- ↑ Пиневич, 2006, с. 199.
Литература[править | править код]
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т.. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп.. — М.: Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0.
- Кассимерис Л., Лингаппа В. Р., Плоппер Д. . Клетки по Льюину. — М.: Лаборатория знаний, 2016. — 1056 с. — ISBN 978-5-906828-23-1.
Источник
ЦИТОПЛАЗМА (ЦП)
Участвуют в спорообразовании.
МЕЗОСОМЫ
При избыточном росте, по сравнению с ростом КС, ЦПМ образует инвагинаты (впячивания) — мезосомы.Мезосомы — центр энергетического метаболизма прокариотической клетки. Мезосомы являются аналогами митохондрий эукариот, но устроены проще.
Хорошо развитые и сложно организованные мезосомы характерны для Грам+ бактерий. У Грам- бактерий мезосомы встречаются реже и просто организованы (в форме петли). Полиморфизм мезосом отмечается даже у одного и того же вида бактерий. У риккетсий мезосомы отсутствуют.
Мезосомы различаются по размеру, форме и локализации в клетке.
По форме различают мезосомы:
– – ламеллярные (пластинчатые),
– – везикулярные (имеющие форму пузырьков),
– – тубулярные (трубчатые),
– – смешанные.
По расположению в клетке различают мезосомы:
– – образующиеся в зоне клеточного деления и формирования поперечной перегородки,
– – к которым прикреплен нуклеоид;
– – сформированные в результате инвагинации периферических участков ЦПМ.
Функции мезосом:
1. Усиливают энергетический метаболизм клеток, так как увеличивают общую «рабочую» поверхность мембран.
2. Участвуют в секреторных процессах (у некоторых Грам+ бактерий).
3. Участвуют вклеточном делении. При размножении нуклеоид движется к мезосоме, получает энергию, удваивается и делится амитозом.
Выявление мезосом:
1. Электронная микроскопия.
Строение.Цитоплазма (протоплазма)—содержимое клетки, окруженное ЦПМ и занимающее основной объем бактериальной клетки. ЦП является внутренней средой клетки и представляет собой сложную коллоидную систему, состоящую из воды (около 75%) и различных органических соединений (белков, РНК и ДНК, липидов, углеводов, минеральных веществ).
Располагающийся под ЦПМ слой протоплазмы более плотный, чем остальная масса в центре клетки. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: нуклеоидом, плазмидами, рибосомами и включениями.
Функции цитоплазмы:
1. Содержит клеточные органеллы.
Выявление цитоплазмы:
1. Электронная микроскопия.
Строение. Нуклеоид— эквивалент ядра эукариот, хотя отличается от него по своей структуре и химическому составу. Нуклеоид не отделен от ЦП ядерной мембраной, не имеет ядрышек и гистонов, содержит одну хромосому, имеет гаплоидный (одиночный) набор генов, не способен к митотическому делению.
Нуклеоид расположен в центре бактериальной клетки, содержит двунитевую молекулу ДНК, небольшое количество РНК и белков. У большинства бактерий двунитевая молекула ДНК диаметром около 2 нм, длиной около 1 м с молекулярной массой 1–3х109 Да замкнута в кольцо и плотно уложена наподобие клубка. У микоплазм молекулярная масса ДНК наименьшая для клеточных организмов (0,4–0,8×109 Да).
ДНК прокариот построена так же, как и у эукариот (рис. 25).
Рис. 25. Строение ДНК прокариот:
А — фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание: 1 — цитозин; 2 — гуанин.
Б — двойная спираль ДНК: Д — дезоксирибоза; Ф — фосфат; А — аденин; Т — тимин; Г — гуанин; Ц — цитозин
Молекула ДНК несет множество отрицательных зарядов, так как каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками — гистонами. В клетках прокариот гистонов нет, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами и ионами Mg2+.
По аналогии с хромосомами эукариот бактериальная ДНК часто обозначается как хромосома. Она представлена в клетке в единственном числе, поскольку бактерии являются гаплоидными. Однако перед делением клетки число нуклеоидов удваивается, а во время деления увеличивается до 4 и более. Поэтому термины «нуклеоид» и «хромосома» не всегда совпадают. При действии на клетки определенных факторов (температуры, pH среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках обнаруживается по одной копии хромосомы.
Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Специальные исследования показали, что хромосомы прокариот — высокоупорядоченная структура. Часть ДНК в этой структуре представлена системой из 20–100 независимо суперспирализованных петель. Суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По периферии нуклеоида располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК). Поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами.
Кроме нуклеоида в цитоплазме бактериальной клетки могут находиться плазмиды — факторы внехромосомной наследственности в виде дополнительных автономных кольцевых молекул двунитевой ДНК с меньшей молекулярной массой. В плазмидах также закодирована наследственная информация, однако она не является жизненно необходимой для бактериальной клетки.
Функции нуклеиода:
1. Хранение и передача наследственной информации, в том числе о синтезе факторов патогенности.
Выявление нуклеоида:
| Рис. 26. Нуклеоид стафилококка (трансмиссивная электронная микроскопия) |
1. Электронная микроскопия: на электронограммах ультратонких срезов нуклеоид имеет вид светлых зон меньшей оптической плотности с фибриллярными, нитевидными структурами ДНК (рис. 26). Несмотря на отсутствие ядерной мембраны, нуклеоид довольно четко отграничен от цитоплазмы.
2. Фазово-контрастная микроскопия нативных препаратов.
3. Световая микроскопия после окраски специфическими для ДНК методами по Фельгену, по Пашкову или по Романовскому-Гимза:
– препарат фиксируют метиловым спиртом;
– на фиксированный препарат наливают краситель Романовского-Гимза (смесь равных частей трех красок — азура, эозина и метиленового синего, растворенных в метаноле) на 24 часа;
– краску сливают, промывают препарат дистиллированной водой, высушивают и микроскопируют: нуклеоид окрашивается в фиолетовый цвет и располагается диффузно в цитоплазме, окрашенной в бледно-розовый цвет.
Источник
Нуклеоти́ды (нуклеозидфосфаты) — группа органических соединений, представляют собой фосфорные эфиры нуклеозидов. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов.
Строение
Нуклеотиды являются сложными эфирами нуклеозидов и фосфорных кислот. Нуклеозиды, в свою очередь, являются N-гликозидами, содержащими гетероциклический фрагмент, связанный через атом азота с C-1 атомом остатка сахара.
В природе наиболее распространены нуклеотиды, являющиеся β-N-гликозидами пуринов или пиримидинов и пентоз — D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды, которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) — соответственно РНК или ДНК[1].
Фосфатный остаток в нуклеотидах обычно образует сложноэфирную связь с 2′-, 3′- или 5′-гидроксильными группами рибонуклеозидов, в случае 2′-дезоксинуклеозидов этерифицируются 3′- или 5′-гидроксильные группы.
Большинство нуклеотидов являются моноэфирами ортофосфорной кислоты, однако известны и диэфиры нуклеотидов, в которых этерифицированы два гидроксильных остатка — например, циклические нуклеотиды циклоаденин- и циклогуанин монофосфаты (цАМФ и цГМФ). Наряду с нуклеотидами — эфирами ортофосфорной кислоты (монофосфатами) в природе также распространены и моно- и диэфиры пирофосфорной кислоты (дифосфаты, например, аденозиндифосфат) и моноэфиры триполифосфорной кислоты (трифосфаты, например, аденозинтрифосфат).
Номенклатура
| Код | Означает | Комплементарная пара |
|---|---|---|
| A | A | T в ДНК; U в РНК |
| C | C | G |
| G | G | C |
| T или U | T в ДНК; U в РНК | A |
| M | A или C | K |
| R | A или G | Y |
| W | A или T | W |
| S | C или G | S |
| Y | C или T | R |
| K | G или T | M |
| V | A или C или G | B |
| H | A или C или T | D |
| D | A или G или T | H |
| B | C или G или T | V |
| X или N | A или C или G или T (U) | любой |
Соединения, состоящие из двух нуклеотидовых молекул, называются динуклеотидами, из трёх — тринуклеотидами, из небольшого числа — олигонуклеотидами, а из многих — полинуклеотидами, или нуклеиновыми кислотами.
Названия нуклеотидов представляют собой аббревиатуры в виде стандартных трёх- или четырёхбуквенных кодов.
Если аббревиатура начинается со строчной буквы «д» (англ. d), значит подразумевается дезоксирибонуклеотид; отсутствие буквы «д» означает рибонуклеотид. Если аббревиатура начинается со строчной буквы «ц» (англ. c), значит речь идёт о циклической форме нуклеотида (например, цАМФ).
Первая прописная буква аббревиатуры указывает на конкретное азотистое основание или группу возможных нуклеиновых оснований, вторая буква — на количество остатков фосфорной кислоты в структуре (М — моно-, Д — ди-, Т — три-), а третья прописная буква — всегда буква Ф («-фосфат»; англ. P).
Латинские и русские коды для нуклеиновых оснований:
- A — А: Аденин;
- G — Г: Гуанин;
- C — Ц: Цитозин;
- T — Т: Тимин (5-метилурацил), не встречается в РНК , занимает место урацила в ДНК;
- U — У: Урацил, встречается у бактериофагов в ДНК, занимает место тимина в РНК.
Общепринятые буквенные коды для обозначения нуклеотидных оснований соответствуют номенклатуре, принятой Международным союзом теоретической и прикладной химии (International Union of Pure and Applied Chemistry, сокращённо — англ. IUPAC, ИЮПАК) и Международным союзом биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology[en], сокращённо — англ. IUBMB). Если при секвенировании последовательности ДНК или РНК возникает сомнение в точности определения того или иного нуклеотида, помимо пяти основных (A, C, T, G, U), используют другие буквы латинского алфавита в зависимости от того, какие наиболее вероятные нуклеотиды могут находиться в данной позиции последовательности. Эти же дополнительные буквы используют для обозначения вырожденных (не совпадающих у разных гомологичных последовательностей) позиций, например при записи последовательности праймеров для ПЦР.
Длину секвенированных участков ДНК (гена, сайта, хромосомы) или всего генома указывают в парах нуклеотидов (пн), или парах оснований (англ. base pairs, сокращённо bp), подразумевая под этим элементарную единицу двухцепочечной молекулы нуклеиновой кислоты, сложенную из двух спаренных комплементарных оснований.
Биологическая роль
1. Универсальный источник энергии (АТФ и его аналоги).
2. Являются активаторами и переносчиками мономеров в клетке (УДФ-глюкоза)
3. Выступают в роли коферментов (ФАД, ФМН, НАД+, НАДФ+)
4. Циклические мононуклеотиды являются вторичными посредниками при действии гормонов и других сигналов (цАМФ, цГМФ).
5. Аллостерические регуляторы активности ферментов.
6. Являются мономерами в составе нуклеиновых кислот, связанные 3′-5′-фосфодиэфирными связями.
История
В 1955 г. Сеймур Бензер для обозначения наименьшего элемента в структуре ДНК, который может быть подвержен спонтанной или индуцированной мутации, ввёл термин мутон, а для обозначения наименьшей единицы рекомбинации — термин рекон. В настоящее время показано, что наименьшей единицей мутации является один нуклеотид (или одно азотистое основание в составе нуклеотида), а рекомбинация возможна между соседними нуклеотидами, поэтому данные термины более не употребляются и имеют лишь историческое значение.
Примечания
См. также
- Ген
- Кодон
- Нуклеотидная последовательность
- Хромосома
Ссылки
- Нуклеотиды — статья из Большой советской энциклопедии.
- Буквенные коды нуклеотидных оснований согласно номенклатуре ИЮПАК [1] и IUBMB[en] [2]
- Пара нуклеотидов (base pair) в Словаре биотехнологических терминов интернет-журнала «Коммерческая биотехнология»
Источник
В отличие от эукариот бактерии не имеют оформленного ядра, однако их ДНК не разбросана по всей клетке, а сосредоточена в компактной структуре, которую называют нуклеоидом. В функциональном отношении он представляет собой функциональный аналог ядерного аппарата.
Что такое нуклеоид
Нуклеоид бактерий — это область в их клетках, содержащая структурированный генетический материал. В отличие от ядра эукариот она не отделена мембраной от остального клеточного содержимого и не имеет постоянной формы. Несмотря на это генетический аппарат бактерий четко отграничен от цитоплазмы.
Сам термин означает “подобный ядру” или “ядерная область”. Впервые эту структуру обнаружил в 1890 г. зоолог Отто Бючли, но ее отличия от генетического аппарата эукариот были выявлены аж в начале 1950-х годов благодаря технологии электронной микроскопии. Название “нуклеоид” соответствует понятию “бактериальная хромосома”, если последняя содержится в клетке в единственном экземпляре.
Нуклеоид не включает в себя плазмиды, которые являются внехромосомными элементами бактериального генома.
Особенности нуклеоида бактерий
Обычно нуклеоид занимает центральный участок бактериальной клетки и ориентирован вдоль ее оси. Объем этого компактного образования не превышает 0,5 мкм3, а молекулярная масса варьирует от 1×109 до 3×109 дальтон. В определенных точках нуклеоид связан с клеточной мембраной.
В состав нуклеоида бактерий входят три компонента:
- ДНК.
- Структурные и регуляторные белки.
- РНК.
ДНК имеет хромосомную организацию, отличную от эукариотической. Чаще всего нуклеоид бактерий содержит одну хромосому или несколько ее копий (при активном росте их количество достигает 8 и более). Этот показатель варьирует в зависимости от вида и стадии жизненного цикла микроорганизма. Некоторые бактерии имеют несколько хромосом с разным набором генов.
В центре нуклеоида ДНК укомплектована достаточно плотно. Эта зона недоступна для рибосом, ферментов репликации и транскрипции. Напротив, дезоксирибонуклеиновые петли периферической области нуклеоида напрямую контактируют с цитоплазмой и представляют собой активные участки бактериального генома.

Количество белкового компонента в нуклеоиде бактерий не превышает 10 %, что примерно в 5 раз меньше, чем в хроматине эукариот. Большая часть белков ассоциирована с ДНК и участвует в ее структурировании. РНК представляет собой продукт транскрипции бактериальных генов, которая осуществляется на периферии нуклеоида.
Генетический аппарат бактерий является динамическим образованием, способным менять свою форму и структурную конформацию. В нем отсутствуют характерные для ядра эукариотической клетки ядрышки и митотический аппарат.
Бактериальная хромосома
В большинстве случаев хромосомы нуклеоида бактерий имеют замкнутую кольцевую форму. Значительно реже встречаются линейные хромосомы. В любом случае эти структуры состоят из одной молекулы ДНК, которая содержит набор генов, необходимых для выживания бактерии.
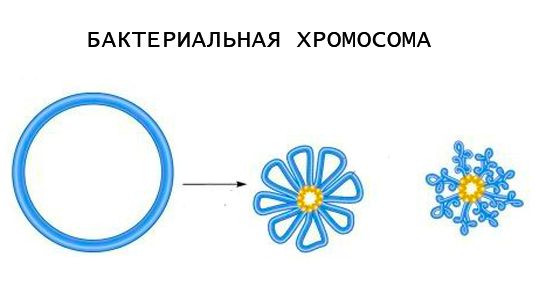
Хромосомная ДНК укомплектована в виде суперспирализованных петель. Количество петель на хромосому варьирует от 12 до 80. Каждая хромосома является полноценным репликоном, так как при удвоении ДНК копируется целиком. Начинается этот процесс всегда из точки начала репликации (OriC), которая прикреплена к плазматической мембране.
Суммарная длина молекулы ДНК в хромосоме на несколько порядков превышает размеры бактерии, поэтому возникает необходимость в ее упаковке, но при сохранении функциональной активности.
В хроматине эукариот эти задачи выполняют основные белки — гистоны. Нуклеоид бактерий имеет в своем составе ДНК-связывающие белки, которые отвечают за структурную организацию генетического материала, а также влияют на экспрессию генов и репликацию ДНК.
К нуклеоид-ассоциированым белкам относятся:
- гистоноподобные белки HU, H-NS, FIS и IHF;
- топоизомеразы;
- белки семейства SMC.
Последние 2 группы оказывают наибольшее влияние на суперспирализацию генетического материала.
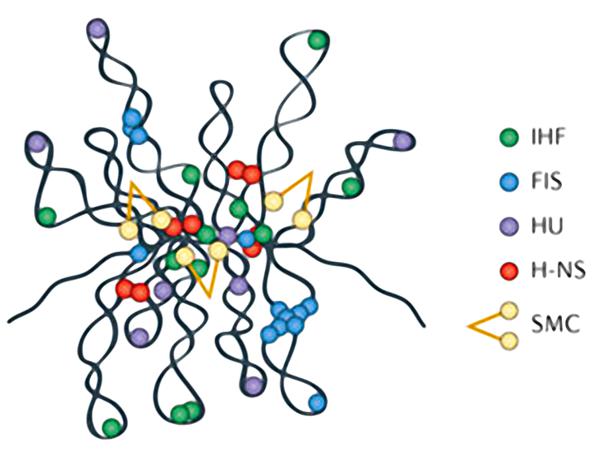
Нейтрализация отрицательных зарядов хромосомной ДНК осуществляется за счет полиаминов и ионов магния.
Биологическая роль нуклеоида
В первую очередь нуклеоид необходим бактериям для того, чтобы хранить и передавать наследственную информацию, а также реализовывать ее на уровне клеточного синтеза. Иными словами, биологическая роль этого образования такая же, как у ДНК.
Другие функции нуклеоида бактерий включают:
- локализацию и компактизацию генетического материала;
- функциональную упаковку ДНК;
- регуляцию метаболизма.
Структурирование ДНК не только позволяет молекуле уместиться в микроскопической клетке, но и создает условия для нормального протекания процессов репликации и транскрипции.
Особенности молекулярной организации нуклеоида создают условия для контроля клеточного метаболизма путем изменения конформации ДНК. Регуляция происходит за счет выпетливания определенных участков хромосомы в цитоплазму, что делает их доступными для ферментов транскрипции, или наоборот, втягивания внутрь.
Способы обнаружения
Существует 3 способа визуального обнаружения нуклеоида в бактериях:
- световая микроскопия;
- фазово-контрастная микроскопия;
- электронная микроскопия.
В зависимости от способа подготовки препарата и метода исследования нуклеоид может выглядеть по разному.
Световая микроскопия
Для выявления нуклеоида при помощи светового микроскопа бактерии предварительно окрашивают таким образом, чтобы нуклеоид имел цвет, отличный от остального клеточного содержимого, — иначе эта структура не будет видна. Также обязательна фиксация бактерий на предметном стекле (при этом микроорганизмы погибают).
Через объектив светового микроскопа нуклеоид выглядит как бобовидное образование с четкими границами, которое занимает центральную часть клетки.
Методы окраски
В большинстве случаев для визуализации нуклеоида методом световой микроскопии используют следующие способы окраски бактерий:
- по Романовскому-Гимзе;
- метод Фельгена.
При окрашивании по Романовскому-Гимзе бактерии предварительно фиксируются на предметном стекле метиловым спиртом, а затем в течение 10-20 минут пропитываются красителем из равной смеси азура, эонина и метиленового синего, растворенных в метаноле. В результате нуклеоид становится фиолетовым, а цитоплазма – бледно-розовой. Перед микроскопией краска сливается, а препарат промывается дистиллятом и высушивается.
В методе Фельгена применяется слабо кислотный гидролиз. В результате освобожденная дезоксирибоза переходит в альдегидную форму и взаимодействует с фуксинсернистой кислотой реактива Шиффа. В итоге нуклеоид становится красным, а цитоплазма приобретает синий цвет.
Фазово-контрастная микроскопия
Фазово-контрастная микроскопия имеет большее разрешение, чем световая. Этот метод не требует фиксации и окраски препарата, — наблюдение происходит за живыми бактериями. Нуклеоид в таких клетках выглядит как светлая овальная область на фоне темной цитоплазмы. Более эффективным метод можно сделать, применив флюоресцентные красители.
Выявление нуклеоида при помощи электронного микроскопа
Существует 2 способа подготовки препарата для исследования нуклеоида под электронным микроскопом:
- ультратонкий срез;
- срез замороженной бактерии.
На электронных микрофотографиях ультратонкого среза бактерии нуклеоид имеет вид состоящей из тонких нитей плотной сетчатой структуры, которая выглядит светлее окружающей цитоплазмы.
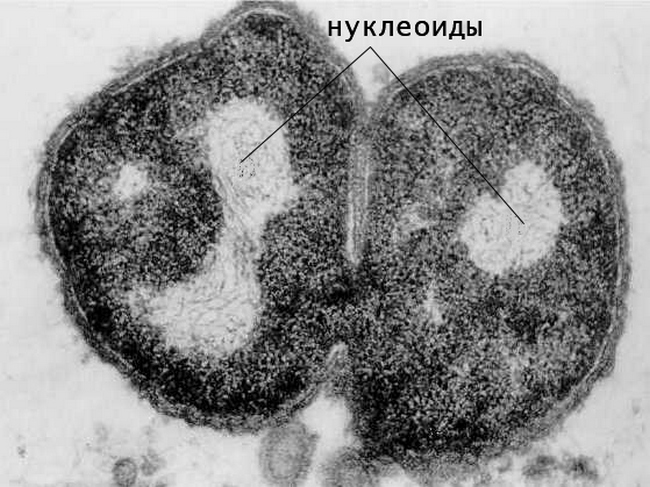
На срезе замороженной бактерии после иммуноокрашивания нуклеоид выглядит как кораллоподобная структура с плотной сердцевиной и тонкими проникающими в цитоплазму выступами.
На электронных фотографиях нуклеоид бактерий чаще всего занимает центральную часть клетки и имеет меньший объем, нежели в живой клетке. Это связано с воздействием химических реактивов, используемых для фиксации препарата.
Источник