
В каких тканях содержится митохондрия

4 июля 2014
Автор
КакПросто!
Гранулы митохондрий были впервые обнаружены в мышечных клетках в 1850 году. Их число в тканях непостоянно. Кроме процентного содержания в клетках, они могут различаться также по размерам, форме и пропорциям.
Инструкция
Митохондрии (от греч. μίτος – нить, χόνδρος – крупинка, зернышко) – это органоиды клетки, участвующие в процессах клеточного дыхания и запасающие энергию в виде молекул АТФ. Именно в форме АТФ энергия становится доступной для энергетических затрат клетки.
Митохондрии есть почти во всех эукариотических клетках, кроме эритроцитов млекопитающих и некоторых паразитических простейших. Количество этих органоидов в клетке может составлять от единиц, как у сперматозоидов, некоторых простейших и водорослей, до многих тысяч. Особенно велико число митохондрий в клетках, нуждающихся в больших энергетических запасах. У животных это мышечные ткани, клетки печени.
Митохондрии обычно имеют шаровидную, овальную или палочковидную формы, но в нейронах, к примеру, они имеют форму нитей, а у некоторых грибов это разветвленные гигантские «энергетические станции».
Несмотря на различие форм, у всех митохондрий принципиально сходный, единый план строения. Подобно пластидам, эти органоиды состоят из двух мембран: внешняя мембрана гладкая, а внутренняя представлена многочисленными складками, перегородками и выступами. Складки внутренней мембраны митохондрий называют кристами. Они имеют большую общую поверхность, и именно на них происходят процессы клеточного окисления.
Как и пластиды в растительных клетках, митохондрии имеют собственный генетический аппарат. Их ДНК, как и у прокариот, представлена кольцевой хромосомой. Это позволяет предположить, что предки митохондрий были свободноживущими безъядерными организмами, которые впоследствии перешли к паразитическому образу жизни или вступили в симбиоз с эукариотами, а затем и вовсе стали неотъемлемой частью их клеток.
Помимо ДНК, митохондрии имеют свою собственную РНК и рибосомы. Перед делением клетки или при интенсивном расходовании ею энергии число митохондрий в результате их деления возрастает, чтобы покрыть растущие (или только предстоящие) потребности клетки в энергии. Если потребность в энергии невелика, число этих органоидов снижается.
Полезный совет
Прокариоты – это безъядерные организмы, не имеющие оформленного ядра, отделенного от цитоплазмы ядерной оболочкой – кариоплазмой. Их единственная кольцевая хромосома расположена непосредственно в цитоплазме клетки, и ее условно называют «бактериальной хромосомой». От ядерных организмов – эукариот – у прокариот есть множество и других принципиальных отличий.
Войти на сайт
или
Забыли пароль?
Еще не зарегистрированы?
This site is protected by reCAPTCHA and the Google
Privacy Policy and
Terms of Service apply.
Источник
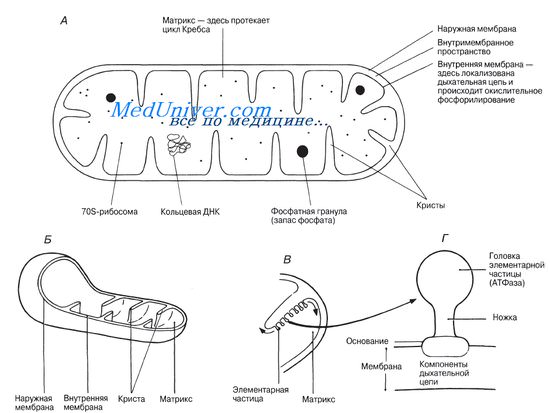
Митохондрии. Строение и функции митохондрий клетки.Митохондрии — органеллы энергообеспечения метаболических процесов в клетке. Размеры их варьируют от 0,5 до 5-7 мкм, количество в клетке составляет от 50 до 1000 и более. В гиалоплазме митохондрии распределены обычно диффузно, однако в специализированных клетках сосредоточены в тех участках, где имеется наибольшая потребность в энергии. Например, в мышечных клетках и симпластах большие количества митохондрий сосредоточены вдоль рабочих элементов — сократительных фибрилл. В клетках, функции которых сопряжены с особо высокими энергозатратами, митохондрии образуют множественные контакты, объединяясь в сеть, или кластеры (кардиомиоциты и симпласты скелетной мышечной ткани). В клетке митохондрии выполняют функцию дыхания. Клеточное дыхание — это последовательность реакций, с помощью которых клетка использует энергию связей органических молекул для синтеза макроэргических соединений типа АТФ. Образующиеся внутри митохондрии молекулы АТФ переносятся наружу, обмениваясь на молекулы АДФ, находящиеся вне митохондрии. В живой клетке митохондрии могут передвигаться с помощью элементов цитоскелета. На ультрамикроскопическом уровне стенка митохондрии состоит из двух мембран — наружной и внутренней. Наружная мембрана имеет относительно ровную поверхность, внутренняя — образует направленные в центр складки, или кристы. Между наружной и внутренней мембранами возникает неширокое (около 15 нм) пространство, которое называется наружной камерой митохондрии; внутренняя мембрана ограничивает внутреннюю камеру. Содержимое наружной и внутренней камер митохондрии различно, и так же, как и сами мембраны, существенно отличается не только по рельефу поверхности, но и по ряду биохимических и функциональных признаков. Наружная мембрана по химическому составу и свойствам близка к другим внутриклеточным мембранам и плазмолемме.
Ее характеризует высокая проницаемость, благодаря наличию гидрофильных белковых каналов. Эта мембрана имеет в своем составе рецепторные комплексы, распознающие и связывающие вещества, поступающие в митохондрию. Ферментный спектр наружной мембраны небогат: это ферменты метаболизма жирных кислот, фосфолипидов, липидов и др. Главной функцией наружной мембраны митохондрии является отграничение органеллы от гиалоплазмы и транспорт необходимых для осуществления клеточного дыхания субстратов. Внутренняя мембрана митохондрий в большинстве клеток тканей различных органов формирует кристы в виде пластин (ламеллярные кристы), что значительно увеличивает площадь поверхности внутренней мембраны. В последней 20-25 % всех белковых молекул составляют ферменты дыхательной цепи и окислительного фосфорилирования. В эндокринных клетках надпочечников и половых желез митохондрии участвуют в синтезе стероидных гормонов. В этих клетках митохондрии имеют кристы в виде трубочек (тубул), упорядоченно расположенных в определенном направлении. Поэтому кристы митохондрий в стероидпродуцирующих клетках названных органов именуются тубулярными. Матрикс митохондрии, или содержимое внутренней камеры, представляет собой гелеобразную структуру, содержащую около 50 % белков. Осмиофильные тельца, описанные при электронной микроскопии, — это резервы кальция. Матрикс содержит ферменты цикла лимонной кислоты, катализирующие окисление жирных кислот, синтез рибосом, ферменты, участвующие в синтезе РНК и ДНК. Общее число ферментов превышает 40. Помимо ферментов, матрикс митохондрии содержит митохондриальную ДНК (митДНК) и митохондриальные рибосомы. Молекула митДНК имеет кольцевидную форму. Возможности внутримитохондриального белкового синтеза ограничены — здесь синтезируются транспортные белки митохондриальных мембран и некоторые ферментные белки, участвующие в фосфорилировании АДФ. Все остальные белки митохондрии кодируются ядерной ДНК, и их синтез осуществляется в гиалоплазме, и в дальнейшем они транспортируются в митохондрию. Жизненный цикл митохондрий в клетке короткий, поэтому природа наделила их двойственной системой воспроизводства — помимо деления материнской митохондрии, возможно образование нескольких дочерних органелл путем почкования. Учебное видео: строение митохондрий и их функции
– Также рекомендуем “Опорно-двигательная система клетки. Промежуточные филаменты клетки.” Оглавление темы “Строение клетки. Клеточные элементы.”: |

Источник
МИТОХОНДРИИ (mitochondria; греч, mitos нить + chondrion зернышко) — органоиды, присутствующие в цитоплазме клеток животных и растительных организмов. Митохондрии принимают участие в процессах дыхания и окислительного фосфорилирования, продуцируют энергию, необходимую для функционирования клетки, представляя, таким образом, ее «силовые станции».
Термин «митохондрии» был предложен в 1894 г. Бендой (С. Benda). В середине 30-х гг. 20 в. удалось впервые выделить М. из клеток печени, что позволило исследовать эти структуры биохим, методами. В 1948 г. Хогебумом (G. Hogeboom) были получены окончательные доказательства того, что М. действительно являются центрами клеточного дыхания. Значительные успехи в изучении этих органоидов были сделаны в 60—70 гг. в связи с применением методов электронной микроскопии и молекулярной биологии.
Рис. 1. Электронограммы различных форм митохондрий из клетки поджелудочной железы крысы: а — удлиненная митохондрия; б — округлая митохондрия; X 38 000.
Форма М. варьирует от почти круглых до сильно вытянутых, имеющих вид нитей (рис. 1), Размер их колеблется от 0,1 до 7 мкм. Количество М. в клетке зависит от типа ткани и функционального состояния организма. Так, в сперматозоидах число М. невелико — ок. 20 (на клетку), в клетках эпителия почечных канальцев млекопитающих их содержится до 300 в каждой, а у гигантской амебы (Chaos chaos) обнаружено 500 000 митохондрий, В одной клетке печени крысы ок. 3000 М., однако в процессе голодания животного число М. может сократиться до 700. Обычно М. распределяются в цитоплазме достаточно равномерно, однако в клетках нек-рых тканей М. могут быть постоянно локализованы в участках, особенно нуждающихся в энергии. Напр., в скелетной мышце М. часто находятся в контакте с контрактильными участками миофибрилл, образуя правильные трехмерные структуры. В сперматозоидах М. образуют спиральный футляр вокруг осевой нити хвоста, что, вероятно, связано с возможностью использовать энергию АТФ, синтезируемую в М., для движений хвоста. В аксонах М. концентрируются вблизи синаптических окончаний, где происходит процесс передачи нервных импульсов, сопровождающийся энергозатратой. В клетках эпителия почечных канальцев М. связаны с выпячиваниями базальной клеточной мембраны. Это вызвано необходимостью постоянного и интенсивного снабжения энергией процесса активного переноса воды и растворенных в ней веществ, протекающего в почках.

Рис. 2. Схема ультраструктуры митохондрии: а —трехмерное изображение митохондрии; б — тонкое строение митохондриальной кристы (соответствует участку, заключенному в рамку на рисунке a): 1 — наружная мембрана; 2 — внутренняя мембрана; 3 — матрикс; 4 — гранулы, содержащие ионы кальция и магния; 5 — кристы; 6 — F1-частицы; 7 — пространство между мембранами; молекулы белка (8) и липидов (9), образующие мембрану; 10 —цепи дыхательных ферментов, расположенные на наружной стороне внутренней мембраны.
Электронно-микроскопически установлено, что М. содержит две мембраны — наружную и внутреннюю. Толщина каждой мембраны ок. 6 нм, расстояние между ними — 6—8 нм. Наружная мембрана гладкая, внутренняя образует сложные выросты (кристы), вдающиеся в полость митохондрии (рис. 2). Внутреннее пространство М. носит название матрикса. Мембраны представляют собой пленку из компактно уложенных молекул белков и липидов, в то время как матрикс подобен гелю и содержит в своем составе растворимые белки, фосфаты и другие хим. соединения. Обычно матрикс выглядит гомогенным, лишь в нек-рых случаях в нем можно обнаружить тонкие нити, трубочки и гранулы, содержащие ионы кальция и магния.
Из особенностей строения внутренней мембраны необходимо отметить наличие в ней сферических частиц ок. 8—10 нм в поперечнике, сидящих на короткой ножке и иногда выступающих в матрикс. Эти частицы были открыты в 1962 г. Фернандес-Мораном (H. Fernandez-Moran). Они состоят из белка, обладающего АТФ-азной активностью, получившего обозначение F1. Белок прикрепляется к внутренней мембране только со стороны, обращенной к матриксу. Частицы F1 располагаются на расстоянии 10 нм друг от друга, а в каждой М. содержится 104—105, таких частиц.
В кристах и внутренних мембранах М. содержится большинство дыхательных ферментов (см.), дыхательные ферменты организованы в компактные ансамбли, распределенные с правильными промежутками в кристах М. на расстоянии 20 нм друг от друга.
М. почти всех типов клеток животных и растений построены по единому принципу, однако возможны отклонения в деталях. Так, кристы могут располагаться не только поперек длинной оси органоида, но и продольно, напр, в М. синаптической зоны аксона. В ряде случаев кристы могут ветвиться. В М. простейших организмов, нек-рых насекомых и в клетках клубочковой зоны надпочечников кристы имеют форму трубочек. Число крист различно; так, в М. клеток печени и половых клеток крист очень мало и они короткие, в то время как матрикс обилен; в М. мышечных клеток кристы многочисленны, а матрикса мало. Существует мнение, что число крист коррелирует с окислительной активностью М.
Во внутренней мембране М. осуществляются параллельно три процесса: окисление субстрата цикла Кребса (см. Трикарбоновых кислот цикл), перенос освободившихся при этом электронов и накопление энергии путем образования макроэргических связей аденозинтрифосфата (см. Аденозинфосфорные кислоты). Основной функцией М. является сопряжение синтеза АТФ (из АДФ и неорганического фосфора) и аэробного процесса окисления (см. Окисление биологическое). Накопленная в молекулах АТФ энергия может трансформироваться в механическую (в мышцах), электрическую (нервная система), осмотическую (почки) и т. д. Процессы аэробного дыхания (см. Окисление биологическое) и сопряженного с ним окислительного фосфорилирования (см.) являются основными функциями М. Кроме того, в наружной мембране М. может происходить окисление жирных к-т, фосфолипидов и нек-рых других соединений.
В 1963 г. Насс и Насс (М. Nass, S. Nass) установили, что в М. содержится ДНК (одна или несколько молекул). Все исследованные до сих пор митохондриальные ДНК из животных клеток состоят из ковалентно замкнутых колец диам. ок. 5 нм. У растений митохондриальная ДНК значительно длиннее и не всегда имеет форму кольца. Митохондриальная ДНК во многих отношениях отличается от ядерной. Репликация ДНК происходит при помощи обычного механизма, однако не совпадает во времени с репликацией ядерной ДНК. Количество генетической информации, заключенной в молекуле митохондриальной ДНК, по-видимому, недостаточно для кодирования всех белков и ферментов, содержащихся в М. Митохондриальные гены кодируют в основном структурные белки мембран и белки, участвующие в морфогенезе митохондрий. М. имеют свои транспортные РНК и синтетазы, содержат все компоненты, необходимые для синтеза белка; их рибосомы меньше цитоплазматических и более похожи на рибосомы бактерий.
Продолжительность жизни М. сравнительно невелика. Так, время обновления половины количества М. составляет для печени 9,6—10,2 сут., для почки — 12,4 сут. Пополнение популяции М. происходит, как правило, из предсуществующих (материнских) М. путем их деления или почкования.
Давно высказывалось предположение, что в процессе эволюции М. возникли, вероятно, путем эндосимбиоза примитивных ядросодержащих клеток с бактериоподобными организмами. Имеется большое число доказательств этому: наличие собственной ДНК, более сходной с ДНК бактерий, чем с ДНК ядра клетки; присутствие в М. рибосом; синтез ДНК-зависимой РНК; чувствительность митохондриальных белков к антибактериальному препарату — хлорамфениколу; сходство с бактериями в реализации дыхательной цепи; морфол., биохим, и физиол, различия между внутренней и наружной мембраной. Согласно симбиотической теории клетка-хозяин рассматривается как анаэробный организм, источником энергии для к-рого является гликолиз (протекающий в цитоплазме). В «симбионте» же реализуется цикл Кребса и дыхательная цепь; он способен к дыханию и окислительному фосфорилированию (см.).
М. являются весьма лабильными внутриклеточными органоидами, раньше других реагирующими на возникновение каких-либо патол, состояний. Возможны изменения числа М. в клетке (вернее, в их популяциях) или изменения их структуры. Напр., при голодании, действии ионизирующего облучения число М. уменьшается. Структурные изменения обычно состоят в набухании всего органоида, просветлении матрикса, разрушении крист, нарушении целостности наружной мембраны.
Набухание сопровождается значительным изменением объема М. В частности, при ишемии миокарда объем М. увеличивается в 10 раз и более. Различают два типа набухания: в одном случае оно связано с изменением осмотического давления внутри клетки, в других случаях — с изменениями клеточного дыхания, сопряженного с ферментативными реакциями и первичными функциональными расстройствами, вызывающими изменения водного обмена. Помимо набухания, может происходить вакуолизация М.
Независимо от причин, вызывающих патол, состояние (гипоксия, гиперфункция, интоксикация), изменения М. довольно стереотипны и неспецифичны.
Наблюдаются такие изменения структуры и функции М., к-рые, по-видимому, становились причиной возникновения болезни. В 1962 г. Луфт (R. Luft) описал случай «митохондриальной болезни». Больному с резко повышенной интенсивностью обмена веществ (при нормальной функции щитовидной железы) была сделана пункция скелетной мышцы и найдено повышенное число М., а также нарушение структуры крист. Дефектные митохондрии в клетках печени наблюдались и при выраженном тиреотоксикозе. Виноград (J. Vinograd) с сотр. (с 1937 по 1969) обнаружил, что у больных с определенными формами лейкемии митохондриальные ДНК из лейкоцитов заметно отличались от нормальных. Они представляли собой открытые кольца или группы сцепленных колец. Частота этих аномальных форм снижалась в результате химиотерапии.
См. также Клетка.
Библиография:
Гаузе Г. Г. Митохондриальная ДНК, М., 1977, библиогр.;
Де Робертис Э., Новинский В. и Саэс Ф. Биология клетки, пер. с англ., М., 1973; Озернюк Н. Д. Рост и воспроизведение митохондрий, М., 1978, библиогр.; Поликар А. и Бесси М. Элементы патологии клетки, пер. с франц., М., 1970; Рудин Д. и Уилки Д. Биогенез митохондрий, пер. с англ., М., 1970, библиогр.; Серов В. В. и Пауков В. С. Ультраструктурная патология, М., 1975; Сэджер Р. Цитоплазматические гены и органеллы, пер. с англ., М., 1975.
Т. А. Залетаева.
Источник